mRNA ポリ(A)テールの不均一性の分析のためのイオン対逆相液体クロマトグラフィー分析法
要約
メッセンジャー RNA(mRNA)バイオポリマーが、新規の治療法として用いられています。SARS-CoV-2 ウイルスに対して有効な mRNA ワクチンの開発は、新規 mRNA 療法の研究を加速させました。有望な mRNA アプリケーションとして、がん治療のためのタンパク質置換療法やワクチン接種のアプローチがあります。mRNA テクノロジーの急速な進歩は、製品の試験のための頑健な分析法次第です。このアプリケーションノートでは、効率的なイオン対逆相液体クロマトグラフィー(IP RP LC)と紫外線(UV)検出を使用する 3' ポリ(A)テール修飾の分析について説明します。この分析法では、RNase T1 による mRNA 分子の消化によってポリ(A)テールを遊離させ、ポリ(A)テールの長さのバリアントの分離向けに最適化したイオン対移動相システムを用います。この分析法は、ポリ(A)テールの不均一性の詳細な分析に適しています。
アプリケーションのメリット
- ポリ(A)テールの不均一性の頑健な分析
- 品質管理試験に適した LC UV 分析法
- 最大 150 塩基の長いオリゴヌクレオチドのポリ(A)テールバリアントの n/n-1 分離
はじめに
治療用メッセンジャー mRNA 分子は 1 本鎖核酸であり、通常は 2,000 ~ 10,000 ヌクレオチドの長さです。治療用 mRNA は、修飾 5' キャップ、3' および 5' の非翻訳領域(UTR)、mRNA コード領域(遺伝子転写配列)、3' 末端のポリ(A)テールなど、いくつかの明確に異なる部分で構成される構造を有します(図 1 を参照)。5' 末端キャップ構造により、細胞内のリボソームを捕捉することで翻訳プロセスが促進されると同時に、RNA の分解が低く抑えられます。3' および 5' の非翻訳領域は、翻訳に直接影響し、多くの場合、RNA 翻訳効率が最大になるように配列が最適化されています。mRNA コード領域は、対象の遺伝子(GOI)をコードします。この部分は目的のタンパク質に翻訳される部分で、多くの場合、クリアランス速度を低下させてタンパク質産生量を増加させるための修飾ヌクレオチド(通常は N1-メチルシュードウリジン)が含まれています。最後に、3' 末端にはポリ(A)テールがあり、RNA の分解を最小限に抑え、mRNA の in-vivo での安定性を維持する上で重要な役割を果たしています。mRNA ワクチンのポリ(A)テール部分の長さは通常 100 ~ 150 ヌクレオチド(nt)です。
SARS-CoV-2 ワクチンの成功により、mRNA 治療の研究活動が加速しています。現在、多数の mRNA 臨床試験が進行中です。mRNA 治療の研究、開発、製造には、頑健な分析法が必要です。イオン対逆相液体クロマトグラフィー(IP RP LC)は、オリゴヌクレオチドの分析に最適な分析法です1-5。 n/n-1 分離は、約 60 nt の長さの分子種まで達成可能であり、これより長いオリゴヌクレオチドや mRNA では次第に難しくなります。mRNA のポリ(A)テールの不均一性の分析は、最先端の分離メソッドや質量分析(MS)を用いても困難な作業です。インタクトな mRNA 分子は多くの場合、分析の前に RNase T1 などの選択的ヌクレアーゼで消化します。RNase T1 は RNA 配列を G ヌクレオチドの 3' 末端側で切断し6、LC MS/MS で分析できる比較的短いオリゴヌクレオチドの混合物が生じます1,5,7。mRNA のポリ(A)テール部分に、切断される G ヌクレオチドは含まれていません。これにより、3' ポリ(A)テールが長さ 100 ~ 150 nt のオリゴヌクレオチドとして mRNA から遊離します(図 1 参照)。ヌクレオチド 1 個分だけ長さが異なる長いオリゴヌクレオチドの分離は、ポリアクリルアミドゲル電気泳動8やキャピラリーゲル電気泳動9, 10、クロマトグラフィー分析法8,9,10,11,13では困難です。
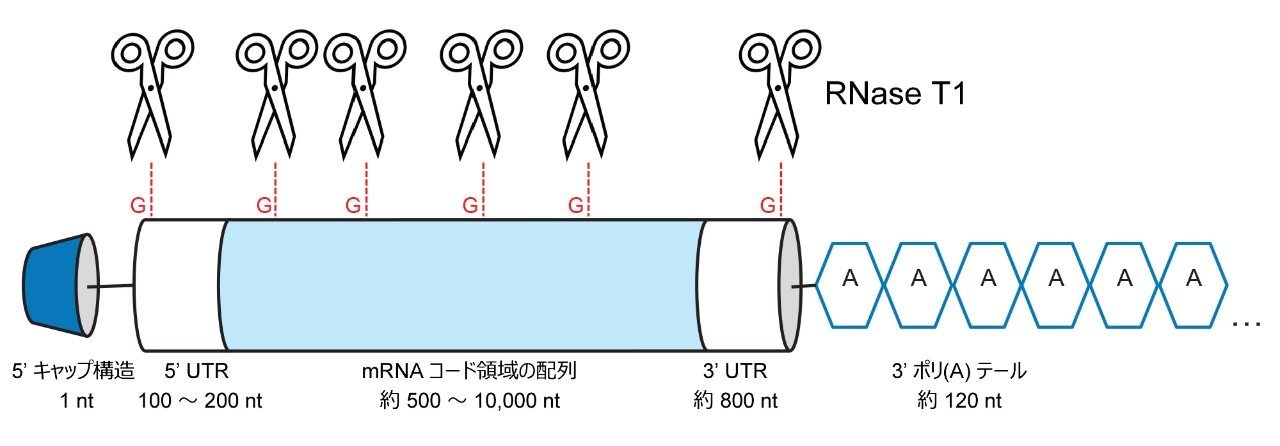 図 1. mRNA 構造の概略図。はさみは、RNase T1 の推定切断部位を示します。消化により短い RNA オリゴヌクレオチドが生成し、3' 末端ポリ(A)テールが遊離します。
図 1. mRNA 構造の概略図。はさみは、RNase T1 の推定切断部位を示します。消化により短い RNA オリゴヌクレオチドが生成し、3' 末端ポリ(A)テールが遊離します。
このアプリケーションノートでは、ポリ(A)テールの不均一性を分析するための高分解能 IP RP LC 分析法について説明します。RNase T1 で遊離したポリ(A)テールは、IP RP LC によって分離し、オリゴリボヌクレオチドアデノシンになります。IP RP LC 分析法での n/n-1 分離は、最大約 150 ヌクレオチドのオリゴヌクレオチドまで可能でした。この分析法は、ポリ(A)テールの不均一性の分析、およびポリ(A)テール中の主要なオリゴヌクレオチド分子種の長さの確認に有用です。また、紫外線検出(SEC UV)を用いるシンプルなサイズ排除クロマトグラフィー分析法を使用して、ポリ(A)テールの平均長を推定することもできます。SEC UV 分析法については、別のアプリケーションノート 720007853 で説明しています。
実験方法
サンプル前処理
EPO mRNA 標準試料は、TriLink Biotechnologies から入手しました(製品番号 L-7209)。EPO mRNA は 5' に Cap 1 構造が含まれ、これはポリアデニル化されており、配列全体に修飾 5-メトキシウリジンが含まれます。Fluc-β mRNA サンプルは AmpTec から入手しました(パーツ番号 M1436/1000-C1-A120-NM-P0)。この mRNA はポリアデニル化されており、配列に非修飾ウリジンが含まれます。mRNA サンプルの 1 mg/mL 溶液を以下のプロトコルを使用して RNase T1 消化しました。このプロトコルでは、50 µL の 1 mg/mL mRNA を 10 µL の rCutSmart 10x バッファー(New England Biolabs、製品番号 B6004S)と混合し、これに 2 µL の RNase T1(Thermo Fisher、製品番号 EN0542、1000 U/µL)を添加しました。サンプルは 37 ℃ で 30 分間消化しました。消化後、1 µL の Quick CIP 酵素(New England Biolabs、製品番号 M0525S、5000 µ/mL)をサンプルに添加して、オリゴヌクレオチドから 3' リン酸基を除きました。脱リン酸化は、室温で 30 分間行いました。このサンプルに 20 µL の 1M TEAA(Sigma、製品番号 90358)および 137 µL の RNase フリー水(Sigma、製品番号 W4502)を添加しました。最終容量約 200 µL のサンプルを、Oasis™ HLB 96 ウェル µElution プレート(Waters™、製品番号 186001828BA)を使用し、50 µL のアセトニトリル、続いて 100 µL の 0.1 M TEAA 溶液でプレートを洗浄するというプロトコルを用いてクリーンアップしました。流速約 0.5 mL/分でサンプルをロードします。サンプルを回収し、2 回目のロードを行います。40 µL の 30% アセトニトリル溶液でプレートからサンプルを溶出します。溶離液 1 ~ 5 µL をイオン対逆相 LC カラムに注入して分析します。100 nt のオリゴリボヌクレオチドアデノシンのカスタム合成は、IDT DNA(米国アイオワ州、Coralville)に依頼しました。
IP RP LC の条件
|
LC システム: |
ACQUITY™ Premier UPLC™(SM-FTN および QSM 搭載) |
|
検出: |
PDA、Ti 5 µL セル、260 nm |
|
カラム: |
ACQUITY Premier Oligonucleotide BEH C18 300 Å、2.1 × 150 mm、1.7 µm(製品番号:186010541) |
|
カラム温度: |
60 ℃ |
|
サンプル温度: |
10 ℃ |
|
注入量: |
1.0 µL(サンプル) |
|
流速: |
0.3 mL/分 |
|
移動相 A: |
100 mM 酢酸オクチルアンモニウム(OAA)含有 40% アセトニトリル/1% ヘキサフルオロイソプロパノール(HFIP)溶液(v:v) |
|
移動相 B: |
100 mM OAA 含有 90% アセトニトリル/1% HFIP 含有 溶液 |
|
移動相 D: |
100% アセトニトリル |
|
グラジエント: |
40 分間で、46% A/54% B/0% D から 28% A/62% B/10% D まで |
|
クロマトグラフィーソフトウェア: |
Empower™ v3.0 |
移動相 A の調製:116.13 g の水と 60.86 g のアセトニトリルを混合し、氷酢酸 1.145 mL、オクチルアミン 3.305 mL、HFIP 2 mL を加えます。よく混ぜ合わせます。
移動相 B の調製:19.355 g の水と 136.92 g のアセトニトリルを混合し、氷酢酸 1.145 mL、オクチルアミン 3.305 mL、HFIP 2 mL を加えます。よく混ぜ合わせます。
結果および考察
IP RP LC 分析法の開発戦略
オリゴヌクレオチドの IP RP LC分離では、イオン対移動相添加剤を使用します1,2,14。 アルキルアミンなどの IP 添加剤は、疎水性 RP 吸着剤に吸着し、正に荷電したアルキルアミンとオリゴヌクレオチドの負に荷電したリン酸基のイオン性相互作用を促進します。IP RP LC では、15 ~ 30 nt の短いオリゴヌクレオチドは容易に分離できますが、40 ~ 60 nt の長いオリゴヌクレオチドの分離はこれより難しくなり、60 ~ 100 nt のオリゴヌクレオチドの分離は非常に難しくなります13。 これは、10 nt のオリゴヌクレオチドと 11 nt のオリゴヌクレオチドでは電荷が 10% 異なりますが(リン酸基がそれぞれ 9 個または 10 個)、100 nt のオリゴヌクレオチドと 101 nt のオリゴヌクレオチドでは違いがわずか 1% であるため、100/101 nt の分離では選択性が低下するためです。次は、長いオリゴヌクレオチドの分離を最適化するために検討した戦略です:(i)効率的な疎水性アルキルアミン(ヘキシルアミン、オクチルアミン)を使用する1,14 、(ii)移動相に高濃度(例:50 ~ 100 mM)の IP 試薬を使用する、(iii)効率のよいカラム(2 ミクロン以下の粒子を充塡した長いカラム)を使用する、(iv)毎分 0.1 ~ 0.5% アセトニトリル等の浅いグラジエントを使用する2。 ACQUITY Premier Oligonucleotide BEH C18 300 Å、2.1 × 150 mm、1.7 µm カラムで、100 mM 酢酸オクチルアンモニウム IP 含有移動相と毎分 0.25% アセトニトリルのグラジエントを組み合わせて使用することで、長さ最大 150 nt のポリ(A)テールオリゴヌクレオチドに最適な分離が得られました。100 mM を超える OAA 濃度は推奨しません。それは、溶出に使用するアセトニトリルの必要濃度が 80% を超えるため、カラムで核酸が析出することがあるためです。一部の市販の mRNA サンプルには、IP RP LC カラムの性能を損なう成分が含まれています。IP RP LC 分析を行う前に、Oasis HLB 96 ウェル µElution プレートを使用してサンプルクリーンアップを行うことを推奨します。
IP RP LC 分析法のポリ(A)テール分析への適用
図 2 は、最適化した IP RP LC 分析法を RNase T1 消化した EPO mRNA(TriLink)および Fluc-β mRNA(AmpTech)の分析に適用したことを示しています。消化によって生成した 2 ~ 30 nt の短いオリゴヌクレオチドは、5 分前に溶出しています(データは示していません)。図 2 では、ポリ(A)分子種が 12 ~ 35 分に溶出しています。長さ 124 nt 付近のピーク群(青色のクロマトグラム)は、EPO mRNA のポリ(A)テールの分析です。赤色のクロマトグラムから、Fluc-β mRNA のポリ(A)テールの方がやや長く、128 nt 付近にピーク群があることが分かります。
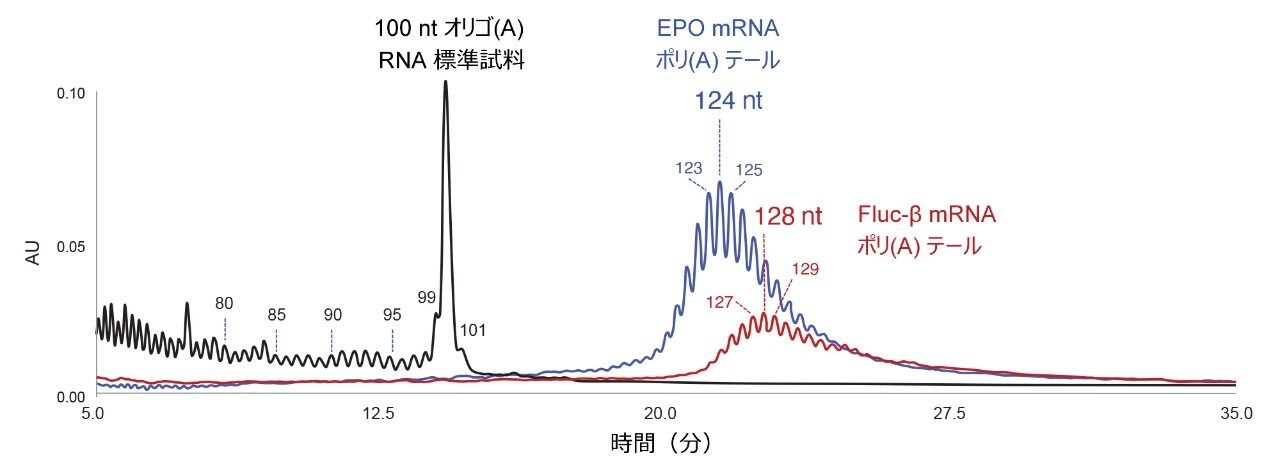 図 2. ACQUITY Premier Oligonucleotide BEH 300 Å 1.7 µm カラムで IP RP LC 分析法を使用した、100 nt のオリゴ(A)合成 RNA オリゴヌクレオチド標準試料(黒色のクロマトグラム)、RNase T1 消化 EPO mRNA(青色のクロマトグラム)および RNase T1 消化 Fluc-β mRNA(赤色のクロマトグラム)の分離。EPO mRNA ポリ(A)テールの一連のピークの中で最も存在量の多いピークは、長さ 124 nt の分子種に対応します。Fluc-β mRNA ポリ(A)テールのピーク群の中で最も存在量の多いピークは、128 nt の分子種に対応します。
図 2. ACQUITY Premier Oligonucleotide BEH 300 Å 1.7 µm カラムで IP RP LC 分析法を使用した、100 nt のオリゴ(A)合成 RNA オリゴヌクレオチド標準試料(黒色のクロマトグラム)、RNase T1 消化 EPO mRNA(青色のクロマトグラム)および RNase T1 消化 Fluc-β mRNA(赤色のクロマトグラム)の分離。EPO mRNA ポリ(A)テールの一連のピークの中で最も存在量の多いピークは、長さ 124 nt の分子種に対応します。Fluc-β mRNA ポリ(A)テールのピーク群の中で最も存在量の多いピークは、128 nt の分子種に対応します。
開発した IP RP LC 分析法は、厳格には MS 検出に対応していません。100 nt オリゴリボヌクレオチドアデノシン標準試料を使用して、最も存在量の多いポリ(A)オリゴヌクレオチドの長さを保持時間から推定しました(黒色のクロマトグラム、図 2)。合成 100 nt オリゴリボヌクレオチド(A)には、完全長生成物から分離した短い n-x 不純物が含まれます。図 2 でラベル付けした 80、85、90、95、99、100、101 nt の分子種について、線形トレンド L(nt)= a × tr + b をプロットしました。保持時間 tr に対する L(nt)の線形キャリブレーションプロットを図 3 に示します。このトレンドを使用して、EPO mRNA および Fluc-β mRNA 消化物中に存在する主なポリ(A)テールのピークの長さ L(nt)を推定しました(図 2 と図 3 を比較)。RNase T1 消化では G の位置の後で切断されるため、遊離したポリ(A)オリゴヌクレオチドの 5' 末端には、1 つまたは複数の追加の C、A、または U ヌクレオチドが含まれる可能性があります。これは、追加の C ヌクレオチドを含む Fluc-β RNA ポリ(A)テールのケースでした。TriLink の製造元は、EPO mRNA の完全配列を提供していません。ポリ(A)テール分子種の長さの正確な長さの割り当てには、mRNA の配列を考慮する必要があります。必要に応じて、RNase T1 消化を RNase A やその他の特異的 RNase で補完することもできます。
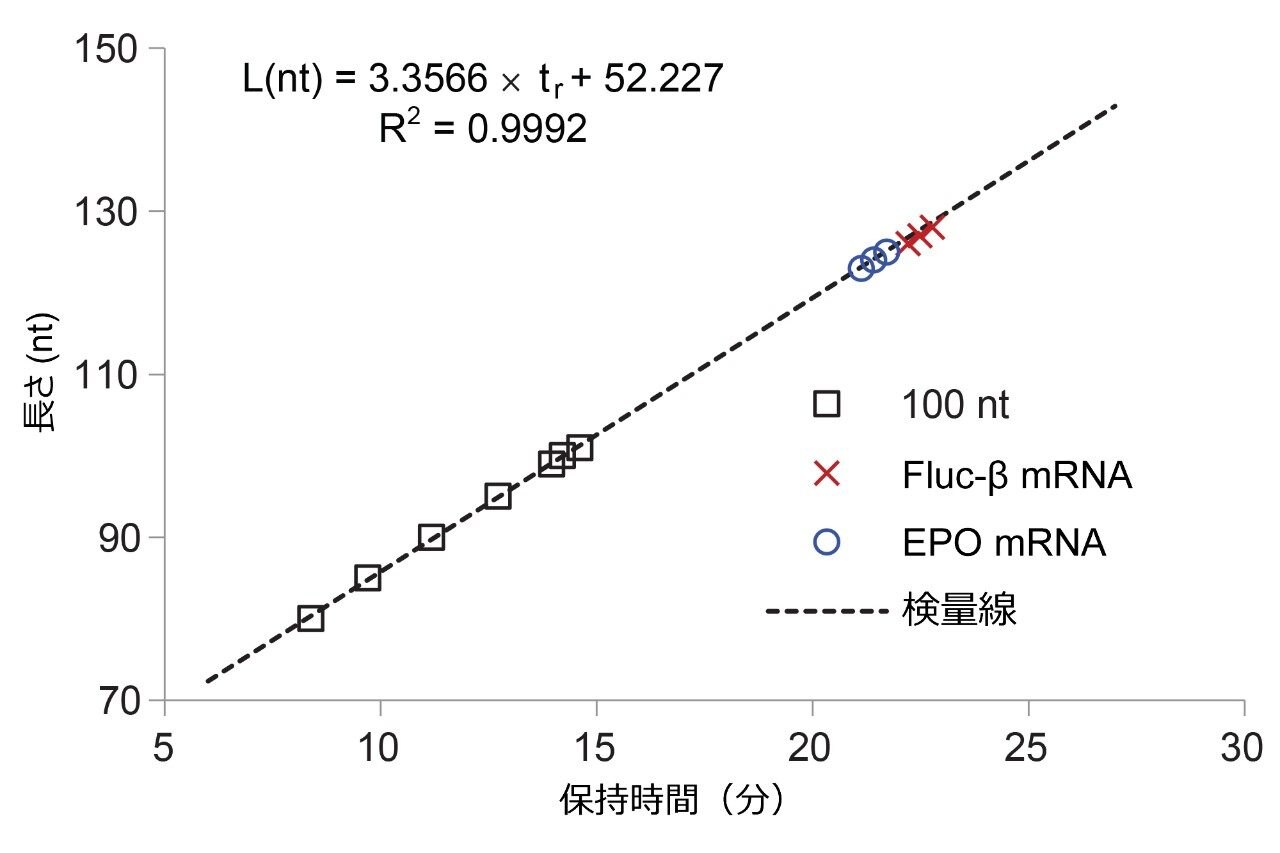 図 3. 検量線 L(nt)= a × tr + b を、100 nt の合成 RNA アデノシン標準試料とその切断合成配列を使用して作成しました(図 2 の黒色のクロマトグラム。80、85、90、95、99、100、101 nt のピーク)。検量線を使用して、mRNA サンプル中で最も存在量の多いポリ(A)テールのピークの長さを計算しました。EPO mRNA の消化物および Fluc-β mRNA の消化物のポリ(A)テールの主要ピークはそれぞれ 124 nt および 128 nt でした。分かりやすくするため、図 2 で検出された最も存在量の多い 3 つのピークのみを図 3 にプロットしました。
図 3. 検量線 L(nt)= a × tr + b を、100 nt の合成 RNA アデノシン標準試料とその切断合成配列を使用して作成しました(図 2 の黒色のクロマトグラム。80、85、90、95、99、100、101 nt のピーク)。検量線を使用して、mRNA サンプル中で最も存在量の多いポリ(A)テールのピークの長さを計算しました。EPO mRNA の消化物および Fluc-β mRNA の消化物のポリ(A)テールの主要ピークはそれぞれ 124 nt および 128 nt でした。分かりやすくするため、図 2 で検出された最も存在量の多い 3 つのピークのみを図 3 にプロットしました。
これら 2 つのポリ A テールサンプルで最も存在量が多い分子種の割り当てを確認するため、ジイソプロピルエチルアミン/HFIP イオン対試薬含有移動相を使用する LC-MS 実験(アプリケーションノート 720007853 で説明されているポリ(A)テールの SEC 測定)を別途実施しました。3 つの分析法(IP RP LC UV、LC-MS、SEC)すべてで、最も存在量の多いポリ(A)テールのピーク(EPO mRNA と Fluc-β mRNA の消化物についてそれぞれ 124 nt ± 1 nt および 128 nt ± 1 nt)に一貫した長さの測定値が得られました。
結論
mRNA のポリ(A)テールの不均一性を分析するために、IP RP LC-UV 分析法を開発しました。最適化した分析法では、ワイドポア ACQUITY Premier Oligonucleotide BEH C18 300 Å、150 × 2.1 mm、1.7 mm カラムと、効率的な酢酸オクチルアンモニウムイオン対形成バッファーを使用します。得られた最大 150 nt の長いオリゴヌクレオチドが分離できる性能は、ポリ(A)テールサンプルの長さおよび不均一性を試験するのに十分です。開発した分析法では、品質管理アッセイとしての使用に適した、シンプルで頑健な UV 検出を使用します。主要なポリ(A)テールオリゴヌクレオチドの長さを、合成オリゴリボヌクレオチド(A)標準試料を用いて作成した検量線により割り当てました。MaxPeak™ High Performance Surfaces を採用した ACQUITY Premier UPLC システムとカラムを使用して、オリゴヌクレオチド分析種について、再現性のよい高い回収率が確保できました4,15-17。
参考文献
- M. Donegan, J.M. Nguyen, and M. Gilar, J. Chromatogr.A, 1666, 462860 (2022).DOI: 10.1016/j.chroma.2022.462860.
- M. Gilar, K.J. Fountain, Y. Budman, U.D. Neue, K.R. Yardley, P.D. Rainville, R.J. Russell, 2nd, and J.C. Gebler, J. Chromatogr.A, 958, 167–182 (2002).
- S.G. Roussis, M. Pearce, and C. Rentel, J. Chromatogr.A, 1594, 105–111 (2019).DOI: 10.1016/j.chroma.2019.02.026.
- G.J. Guimaraes, J.M. Sutton, M. Gilar, M. Donegan, and M.G. Bartlett, J. Pharm.Biomed.Anal., 208, 114439 (2022).DOI: 10.1016/j.jpba.2021.114439.
- A. Goyon, P. Yehl, and K. Zhang, J. Pharm.Biomed.Anal., 182, 113105 (2020).DOI: 10.1016/j.jpba.2020.113105.
- T. Jiang, N. Yu, J. Kim, J.R. Murgo, M. Kissai, K. Ravichandran, E.J. Miracco, V. Presnyak, and S. Hua, Anal.Chem., 91, 8500–8506 (2019).DOI: 10.1021/acs.analchem.9b01664.
- V.B. Ivleva, Y.Q. Yu, and M. Gilar, Rapid Commun Mass Spectrom, 24, 2631–2640 (2010).DOI: 10.1002/rcm.4683.
- L.T. França, E. Carrilho, and T.B. Kist, Q. Rev. Biophys., 35, 169–200 (2002).DOI: 10.1017/s0033583502003797.
- E. Carrilho, M.C. Ruiz-Martinez, J. Berka, I. Smirnov, W. Goetzinger, A.W. Miller, D. Brady, and B.L. Karger, Anal.Chem., 68, 3305–3313.(1996).
- M. Gilar, A. Belenky, Y. Budman, D.L. Smisek, and A.S. Cohen, J Chromatogr B Biomed Sci Appl, 714, 13–20 (1998).
- J.R. Thayer, Thermo Scientific Application note 21996, (2014).
- C.G. Huber, E. Stimpf, P.J. Oefner, and G.K. Bonn, LC-GC, 14, 114–127 (1996).
- M. Gilar and U.D. Neue, J. Chromatogr.A, 1169, 139–150 (2007).DOI: 10.1016/j.chroma.2007.09.005.
- S.M. McCarthy, M. Gilar, and J. Gebler, Anal Biochem, 390, 181-188 (2009).DOI: 10.1016/j.ab.2009.03.042.
- J.M. Nguyen, M. Gilar, B. Koshel, M. Donegan, J. MacLean, Z. Li, and M.A. Lauber, Bioanalysis, (2021).DOI: 10.4155/bio-2021–0115.
- M. Gilar, M. DeLano, and F. Gritti, J. Chromatogr.A, 1650, 462247 (2021).DOI: 10.1016/j.chroma.2021.462247.
- M. DeLano, T.H. Walter, M.A. Lauber, M. Gilar, M.C. Jung, J.M. Nguyen, C. Boissel, A.V. Patel, A. Bates-Harrison, and K.D. Wyndham, Anal.Chem., 93, 5773–5781 (2021).DOI: 10.1021/acs.analchem.0c05203.
720007873JA、2023 年 2 月