Ion-Pairing Reversed Phase LC-MS Analysis of Poly(A) Tail Heterogeneity Using the BioAccord LC-MS System
Abstract
This application note demonstrates the advantages of the BioAccord LC-MS System for the analysis of Poly(A) Tail mRNA heterogeneity.
Benefits
- An LC-MS assay developed on the BioAccord LC-MS System running under waters_connect informatics platform enables robust analysis of mRNA Poly(A) Tail length and heterogeneity, including average molecular weight information for the Poly(A) Tail oligonucleotide region common to therapeutic and prophylactic mRNAs
Introduction
Messenger RNAs (mRNAs) have recently emerged as a new class of biopharmaceuticals, as demonstrated by the successful development and approval of two highly efficacious vaccines based on the mRNA sequence encoding for the SARS-CoV-2 spike protein.1–2 The development of analytical methods for monitoring the critical attributes of mRNA molecules is essential for proper control of the manufacturing process, but the development of such methods for the analysis of large mRNAs (>1,000 nucleotides), including mRNA vaccines, is challenging.
One of the functional structures common to all therapeutic mRNAs is the Poly(A) Tail, a long (50–250 nucleotides) sequence at the 3’-end of the molecule that contains only adenosine residues.3–6 The 3’-end oligonucleotide is typically isolated from the mRNA molecule for analysis using the RNase T1 ribonuclease enzyme, which cleaves after guanosine residues.7–8
Here we investigated the capabilities of the BioAccord LC-MS System for intact mass analysis of a Poly(A) Tail oligonucleotide fragment.
Experimental
Reagents and Sample Preparation
N,N-diisopropylethylamine (DIPEA, 99.5% purity, catalogue number 387649-100ML), 1,1,1,3,3,3-hexafluoro-2-propanol (HFIP, 99% purity, catalogue number 105228-100G), ethanol (HPLC grade, catalogue number 459828-2L), and ammonium bicarbonate (LiChropur LC-MS Supelco reagent, catalogue number 5330050050) were purchased from Millipore Sigma (St Louis, MO). Acetonitrile (LC-MS grade, catalogue number 34881-1L) and methanol (LC-MS grade, catalogue number 34966-1L) were obtained from Honeywell (Charlotte, NC). HPLC grade Type I deionized (DI) water was purified using a MilliQ system (Millipore, Bedford, MA). Mobile phases were prepared fresh daily.
Ultrapure nuclease-free water (catalogue no J71786.AE) for mRNA digestions was purchased from Thermo Fisher Scientific (Waltham, MA).
Twenty nanomoles of a 120-mer polyadenosine mRNA oligonucleotide (Poly A Tail) were purchased from Integrated DNA Technologies (Coralville, IA). The oligonucleotide sequence contained a cytidine incorporated at position 28, a uridine incorporated at position 56, a guanosine incorporated at position 84 and another uridine at position 112 as indicated by the sequence 5’- AAA AAA AAA AAA AAA AAA AAA AAA AAA CAA AAA AAA AAA AAA AAA AAA AAA AAA AUA AAA AAA AAA AAA AAA AAA AAA AAA AAG AAA AAA AAA AAA AAA AAA AAA AAA AAA UAA AAA AAA -3’. These four residues were inserted in the Poly A Tail sequence in order to make the 120-mer RNA nucleotide more stable during chemical synthesis. The full length 120-mer oligo product (FLP) was not further purified by HPLC in order to prevent its potential degradation during a large scale chromatographic purification process. The elemental composition of the synthetic Poly A Tail oligonucleotide is C1197 H1439 N592 O724 P119 and its average molecular weight is 39,388.64 Da.
Stock solutions of the synthetic Poly A Tail oligonucleotide were prepared in DI water at a concentration of 10 µM, from which a 5 µL volume was injected, which corresponds to loading 50 picomoles of the 120-mer oligonucleotide on-column.
Firefly luciferase mRNA (Fluc-beta mRNA, catalogue no M1436/1000-C1-A120-NM-PO, with a concentration of 1.63 mg/mL) was obtained from Amp Tec (Hamburg, Germany). The mRNA was digested with RNase T1 ribonuclease from Aspergillus oryzae (purchased from Millipore Sigma: catalogue no R1003-100KU). For mRNA digestion, 20 µL of Fluc mRNA were mixed with 5 µL digestion buffer (100 mM ammonium bicarbonate), 13 µL of nuclease-free water and 2 µL of a 1:100 diluted solution of the RNase T1 ammonium sulfate solution (prepared also in nuclease-free water). The digestion mixture was prepared in a QuanRecovery MaxPeak 300 µL vial and incubated at 37 oC for 15 min, then immediately analyzed by LC-MS.
All datasets were acquired in UNIFI App ver. 2.1.2.4 and processed using the INTACT Mass App ver 1.6.0.18 within the waters_connect informatics platform.
The MaxEnt 1 charge deconvolution algorithm was used for processing of oligonucleotide ESI-MS spectra to produce accurate intact mass measurements.
LC Conditions
|
LC-MS system: |
BioAccord LC-MS System with ACQUITY Premier UPLC (Binary) |
|
Pre-Column: |
VanGuard FIT cartridge holder (p/n: 186007949) containing a 2.1 x 5 mm ACQUITY Premier FIT cartridge packed with 1.7 µm BEH C18 particles (p/n: 186009459) |
|
Column: |
ACQUITY Premier OST Column 1.7 µm, 130 Å, 2.1 x 50 mm, (p/n: 186009484) |
|
Column temperature: |
60 oC |
|
Flow rate: |
300 µL/min |
|
Mobile phases: |
Solvent A: 8 mM DIPEA (N,N-diisopropylethylamine), 40 mM HFIP (1,1,1,3,3,3-hexafluoroisopropanol), in DI water, pH 8.8 Solvent B: 4 mM DIPEA, 4 mM HFIP in 75% ethanol |
|
Sample temperature: |
6 oC |
|
Sample vials: |
QuanRecovery MaxPeak HPS vials (p/n: 186009186) |
|
Injection volume: |
5 µL |
Wash Solvents
|
Purge solvent: |
50% MeOH |
|
Sample manager wash solvent: |
50% MeOH |
|
Seal wash: |
20% acetonitrile in DI water |
Gradient Table
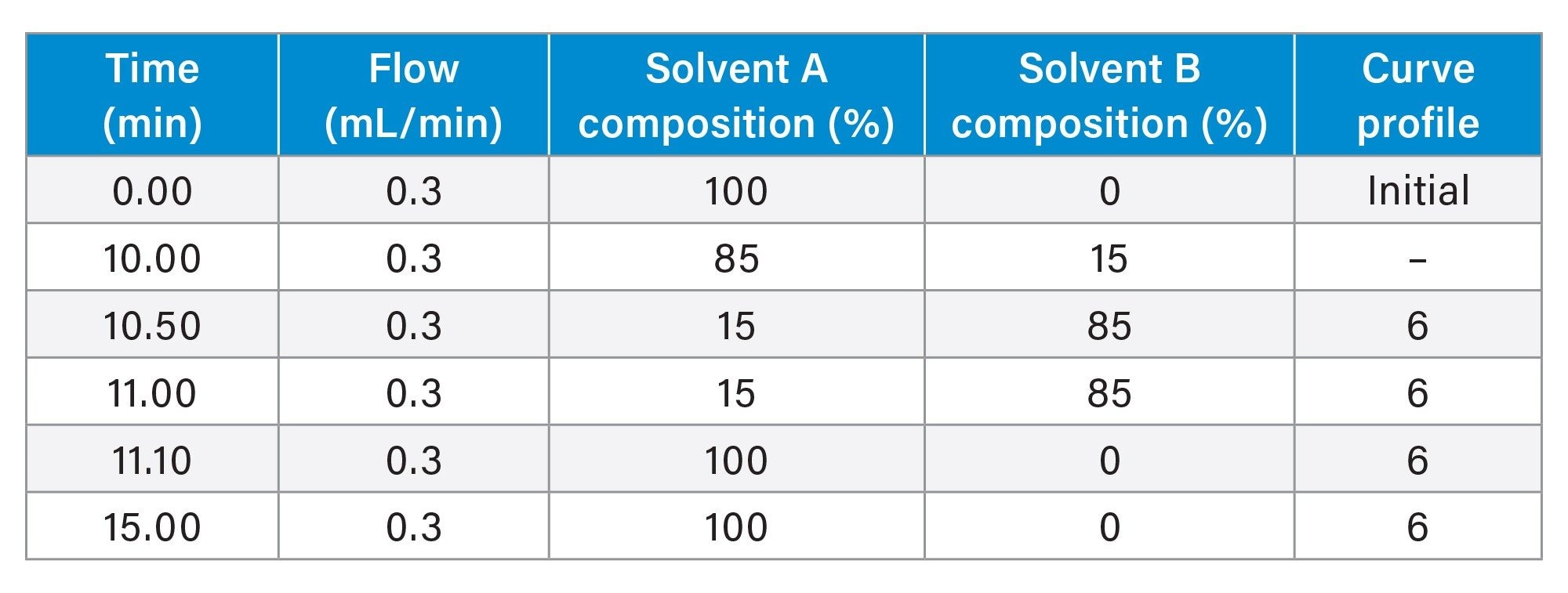
MS Conditions
|
Acquisition mode: |
Full scan |
|
Ionization mode: |
ESI(-) |
|
Capillary voltage: |
0.8 kV |
|
Cone voltage: |
45 V |
|
Source temperature: |
120 oC |
|
Desolvation temperature: |
500 oC |
|
Desolvation gas (N2) pressure: |
6.5 bar |
|
TOF mass range: |
400–5000 |
|
Acquisition rate: |
2 Hz |
|
Lock-mass: |
waters_connect lockmass solution (p/n: 186009298) |
|
Informatics platform for data acquisition and processing: |
waters_connect |
|
Data acquisition: |
UNIFI App ver 2.1.2.4 |
|
Data processing: |
INTACT Mass App ver 1.6.0.18 |
Results and Discussion
Since its discovery in 1961, mRNA has been proposed as therapeutic modality for treatment of a variety of diseases.2 Several structural elements are common to all mRNA molecules designed for therapeutic and prophylactic use, including the Cap sequence located at the 5’-end of the molecule, two untranslated regions, the open reading frame sequence (ORF) - encoding for the gene of interest and the Poly(A) tail located at the 3’-end of the molecule. These structural elements are depicted in a cartoon shown as Figure 1. Similar in structure to a naturally occurring cellular mRNA, a synthetic mRNA is engineered to contain a modified Cap sequence (~50 nucleotides)9 and a long chain of adenosines (50–250 nt) for the Poly(A) tail.3–6 These two modified regions are critical quality attributes (CQAs) of therapeutics mRNAs, playing an important role in mRNA recognition by the cellular machinery responsible for in vitro transcription (IVT). In addition, the 5’-Cap and the 3’-Poly(A) tail improve the efficiency of the IVT process.1 Finally, because the Cap and the Poly(A) regions are located at the termini of the mRNA molecule, they play critical roles for stability of synthetic mRNAs.
 Figure 1. Schematic diagram showing the structural elements of an mRNA molecule. The Poly(A) Tail oligonucleotide, located at the 3’-termini, is a typical cleavage product generated by treatment with RNAse T1 ribonuclease.
Figure 1. Schematic diagram showing the structural elements of an mRNA molecule. The Poly(A) Tail oligonucleotide, located at the 3’-termini, is a typical cleavage product generated by treatment with RNAse T1 ribonuclease.
LC-MS Method Optimization for a Synthetic Poly(A) Tail Oligonucleotide
Several LC-MS assays have been previously reported for the analysis of intact 5’-Cap oligonucleotides as well as intact Poly(A) tail variants.10–13,14,15 Compared to the analysis of 5’-Cap oligonucleotides, the LC-MS characterization of Poly(A) tails is more challenging for several reasons. The Poly(A) tail contains a longer nucleotide sequence, therefore in negative ESI-MS studies it is more difficult to ionize relative to the much shorter Cap sequence. Also, the Poly(A) tail exhibits higher heterogeneity and reduced stability after preparation, being highly prone to enzymatic degradation. In addition, the biological processes responsible for synthesis of the Poly(A) tails in different species are not fully understood, therefore it is difficult to predict exactly all the different sequence variants that might be produced following the IVT process.3–6 For these reasons, for the initial experiments, we decided to use a chemically synthesized analogue of a Poly(A) tail because the oligo sequence could be controlled better and the 120-mer oligonucleotide can be engineered to be more stable after solubilization in DI water, allowing for long-term LC-MS studies (up to four weeks after sample solubilization).
The synthetic Poly(A) tail was first analyzed using an ion-pairing mobile phase suitable for the separation of shorter oligonucleotides, such as resolving siRNA oligos (typically 20 to 25 nucleotides) and their impurities.16–18 The UV chromatogram showing the separation of the crude (unpurified) synthetic 120-mer Poly(A) oligomer is displayed in Figure 2. The TEA (triethylamine) ion pairing reagent is clearly not suitable for separation of such larger oligos as demonstrated by the broad (~0.3 min wide) chromatographic peak shown in this figure.
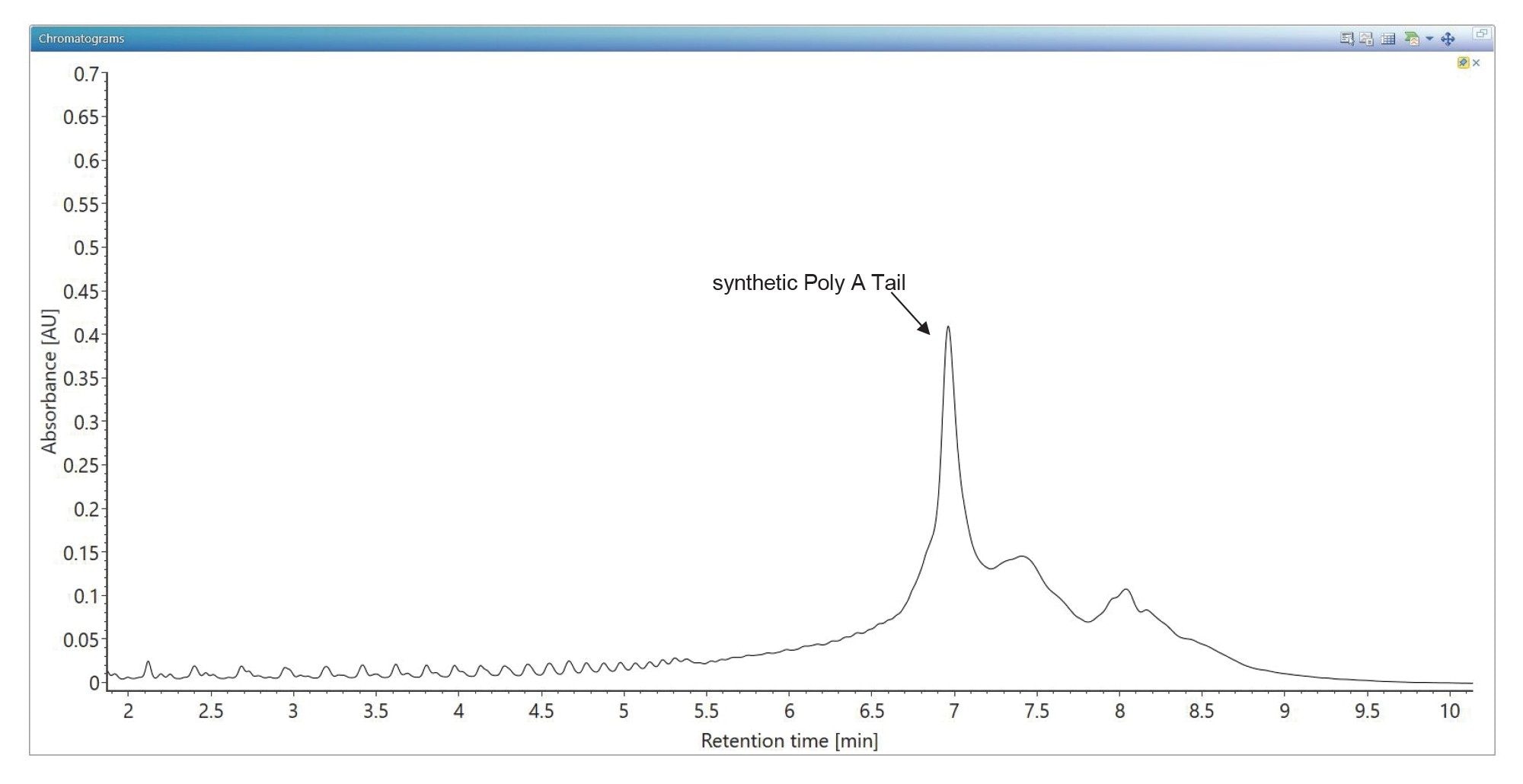 Figure 2. TUV chromatogram showing the IP-RP separation of the 120-mer synthetic Poly(A) Tail oligonucleotide using TEA (triethylamine) as the ion pairing reagent. Solvent A contained 7 mM TEA and 40 mM HFIP in deionized water, while Solvent B contained 3.5 mM TEA, 20 mM HFIP in 50% methanol. TEA is not suitable for separation of large oligos (>40-mers) as it produces rather broad chromatographic peaks (0.3 min peak width) for these species.
Figure 2. TUV chromatogram showing the IP-RP separation of the 120-mer synthetic Poly(A) Tail oligonucleotide using TEA (triethylamine) as the ion pairing reagent. Solvent A contained 7 mM TEA and 40 mM HFIP in deionized water, while Solvent B contained 3.5 mM TEA, 20 mM HFIP in 50% methanol. TEA is not suitable for separation of large oligos (>40-mers) as it produces rather broad chromatographic peaks (0.3 min peak width) for these species.
A branched ion pairing reagent – DIPEA (N,N-diisopropylethylamine) is recommended for separation of large oligos, including single guide RNA oligos (sgRNAs) that are typically 100-mers.7,19 As shown by the overlaid UV chromatograms shown in Figure 3, the chromatographic shape improves considerably (peak width ~0.1 min) for the Poly(A) Tail in the presence of DIPEA. Three different organic solvents were investigated as mobile phase B eluents, including methanol, acetonitrile and ethanol, in order to optimize the intensity of the ESI-MS spectrum for the 120-mer oligo. Methanol is a commonly used solvent for TEA separation of short oligos, but in the case of large oligos, the other two organic solvents (ACN and EtOH) provide slightly sharper Poly(A) Tail peaks, as indicated by chromatograms in Figure 3.16–18
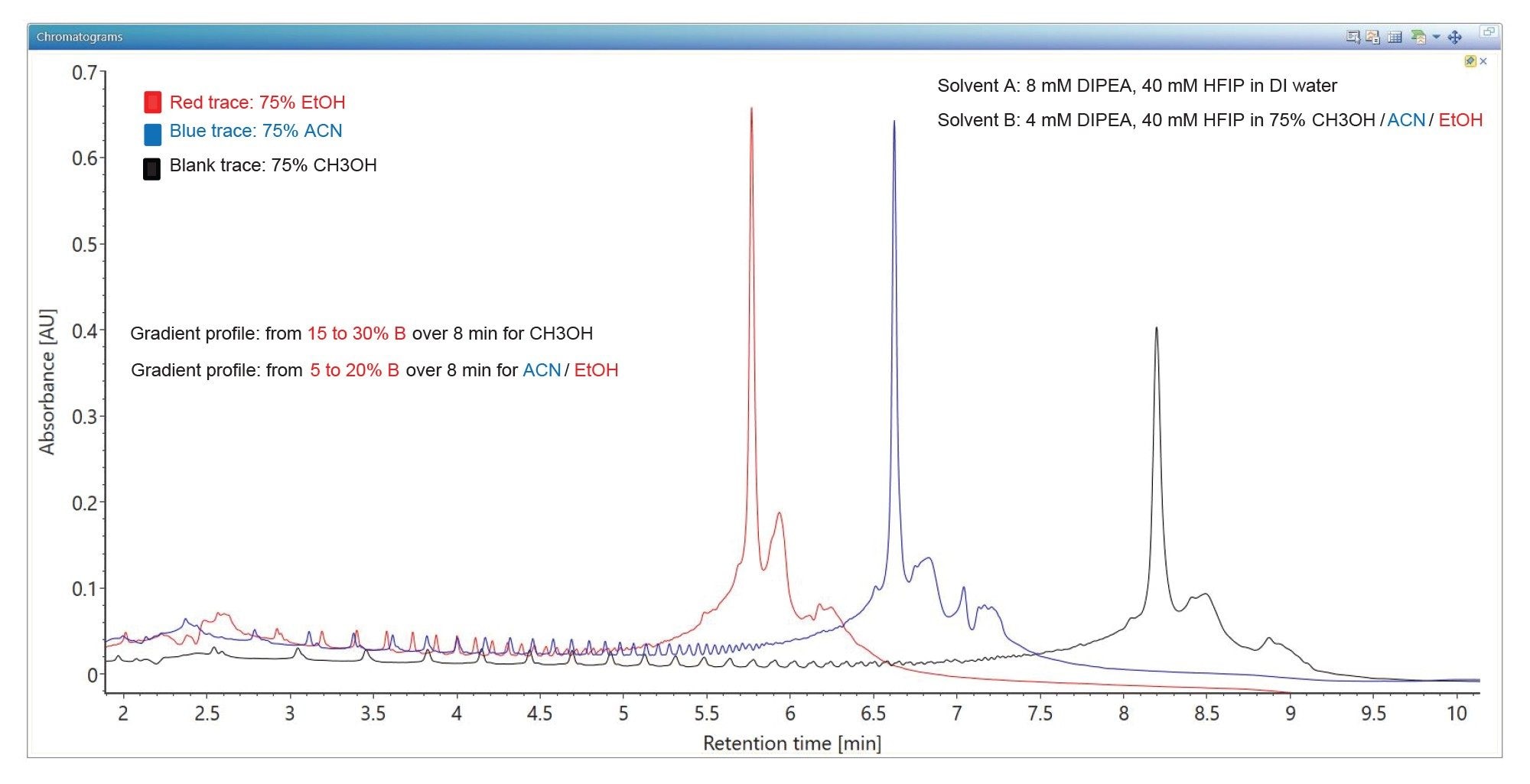 Figure 3. Overlaid TUV chromatogram showing the IP-RP separation of the 120-mer synthetic Poly(A) Tail oligonucleotide using DIPEA (N,N-diisopropylethylamine) as the ion pairing reagent in the presence of three different organic solvents. Solvent A contained 8 mM DIPEA and 40 mM HFIP in deionized water, while Solvent B contained 4 mM DIPEA, 4 mM HFIP in 75% organic solvent, including: 1) methanol (black trace); 2) acetonitrile (blue trace) and 3) ethanol (red trace). DIPEA perform much better compared to TEA, producing sharper chromatographic peaks (~0.1 min peak width) for large oligos like the Poly(A) tail 120-mer oligonucleotide.
Figure 3. Overlaid TUV chromatogram showing the IP-RP separation of the 120-mer synthetic Poly(A) Tail oligonucleotide using DIPEA (N,N-diisopropylethylamine) as the ion pairing reagent in the presence of three different organic solvents. Solvent A contained 8 mM DIPEA and 40 mM HFIP in deionized water, while Solvent B contained 4 mM DIPEA, 4 mM HFIP in 75% organic solvent, including: 1) methanol (black trace); 2) acetonitrile (blue trace) and 3) ethanol (red trace). DIPEA perform much better compared to TEA, producing sharper chromatographic peaks (~0.1 min peak width) for large oligos like the Poly(A) tail 120-mer oligonucleotide.
While methanol provided ~3 fold increase in ESI-MS signal intensity in the presence of DIPEA vs TEA (data not shown), the acetonitrile and ethanol further improved Poly(A) Tail oligonucleotide MS response, as demonstrated by the combined ESI-MS spectra displayed in Figure 4. Acetonitrile is typically the preferred organic solvent for IP/RP LC-MS of large oligos, but ethanol was recently proposed as an alternative for Poly(A) Tail analysis.7,15,19 The combined ESI-MS spectra presented in Figure 4 confirms that ethanol can provide an improved ESI-MS signal over acetonitrile.
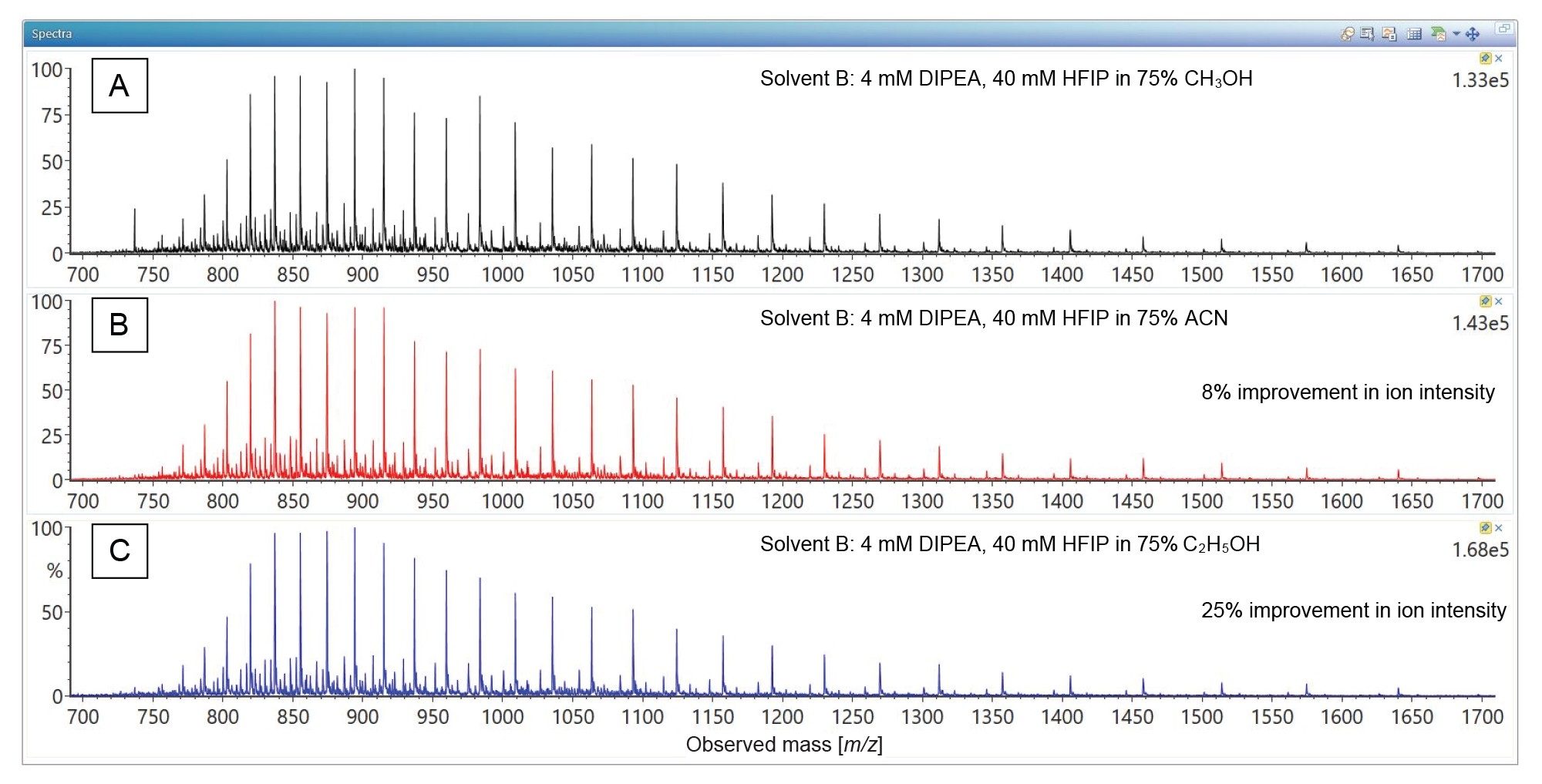 Figure 4. Combined ESI-MS spectra recorded for the Poly(A) Tail oligonucleotide following the IP-RP separations displayed in Figure 4. Solvent A contained 8 mM DIPEA and 40 mM HFIP in deionized water, while Solvent B contained 4 mM DIPEA, 4 mM HFIP in 75% organic solvent, including: methanol (panel A), acetonitrile (panel B), and ethanol (panel C). Ethanol was the preferred solvent since it produced the strongest ESI-MS signal.
Figure 4. Combined ESI-MS spectra recorded for the Poly(A) Tail oligonucleotide following the IP-RP separations displayed in Figure 4. Solvent A contained 8 mM DIPEA and 40 mM HFIP in deionized water, while Solvent B contained 4 mM DIPEA, 4 mM HFIP in 75% organic solvent, including: methanol (panel A), acetonitrile (panel B), and ethanol (panel C). Ethanol was the preferred solvent since it produced the strongest ESI-MS signal.
The intact mass ESI-MS spectrum recorded using 8 mM DIPEA 40 mM HFIP in 75% ethanol as eluent B is displayed (Figure 5B) and the deconvolved spectrum is shown in the panel above (Figure 5A). The ESI-source parameters (ESI voltage, cone voltage, source temperature, desolvation temperature) were optimized to increase the sensitivity of the ESI-MS signal and minimize the levels of sodium adducts. It is also worth mentioning that the choice of the ion pairing reagent, the amount and chemical purity of all the other reagents, as well as the mobile phase preparation procedures are critical in generating cleaner, low-adduct ESI-MS spectra which facilitates the sensitive detection of lower-abundance oligonucleotide species.
In addition to the main target of the chemical synthesis - the 120-mer oligonucleotide, the crude sample contains some shorter length oligonucleotide impurities clearly visible, eluting before the major peak in the chromatograms presented in Figures 2 and 3. It is possible that slightly longer or shorter Poly(A) species, exceeding or being just under the 120-mer target, are present in the crude sample as well, but the RP-IP chromatography does not have the capability to resolve such closely related species. However, it has been shown that size-exclusion chromatography (SEC) is able to better resolve these types of mixtures.20
For co-eluting oligonucleotides, the waters_connect INTACT Mass application performs automatic deconvolution of all detected oligonucleotides (above a user specified intensity threshold) regardless of their retention times or chromatographic profile.19,21 This approach (automated deconvolution, non-RT based) is especially well suited for the analysis of co-eluting oligonucleotide impurities distributed around a parent oligonucleotide species. All the ESI-MS spectra belonging to oligonucleotides eluting between 5.5 and 6.5 min in the red trace chromatogram shown in Figure 3 were automatically deconvolved using the MaxEnt1 algorithm, with the software self-optimizing the deconvolution settings.22
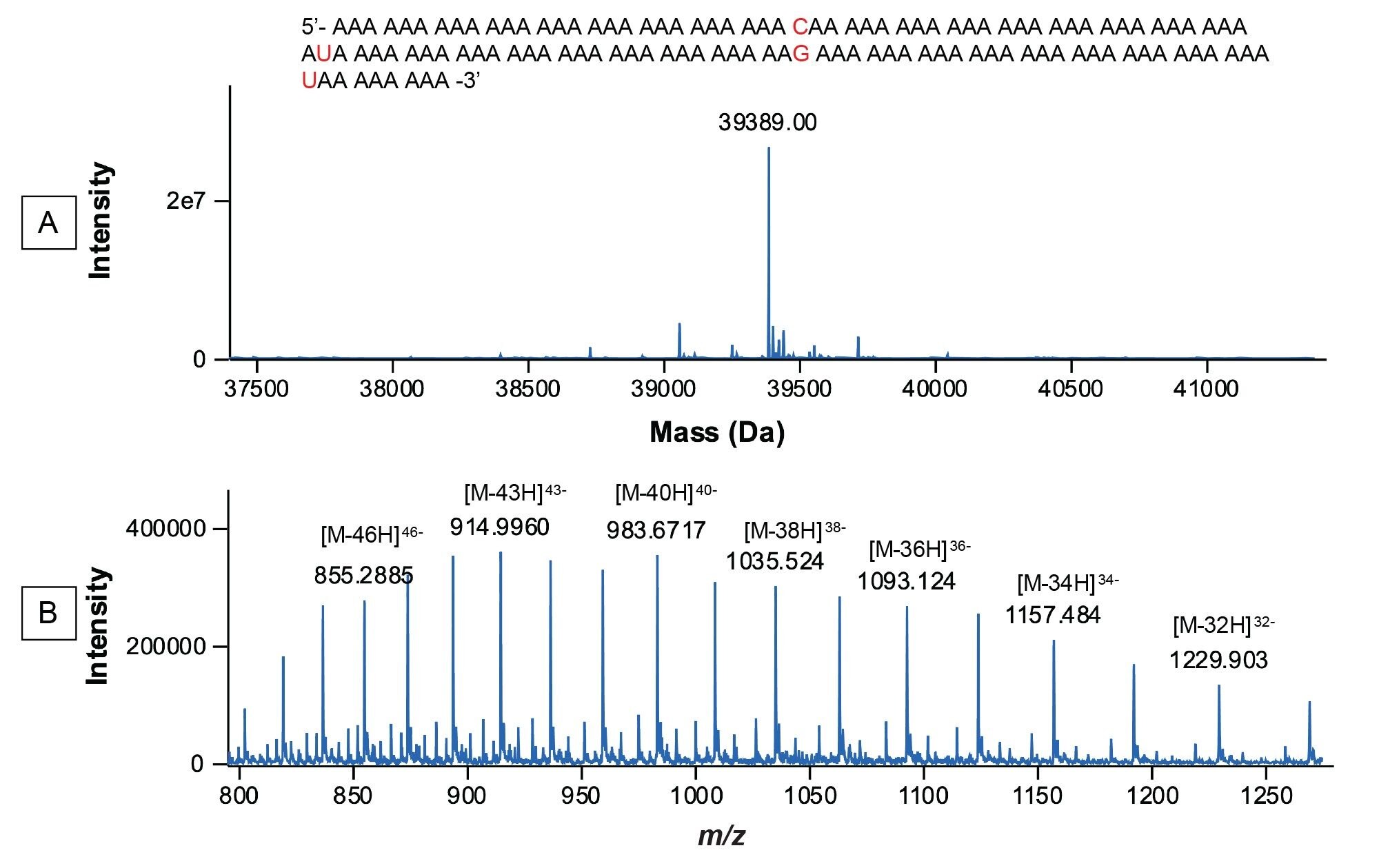 Figure 5. INTACT Mass spectra recorded for the synthetic 120-mer Poly(A) Tail oligonucleotide: (A) MaxEnt1 charge deconvolved spectrum; (B) raw ESI-MS spectrum. The sequence of the 120-mer shown in panel A contains a cytidine incorporated at position 28, a uridine incorporated at position 56, a guanosine incorporated at position 84, and another uridine at position 112. These four residues are used for increasing the chemical stability of the Poly(A) Tail.
Figure 5. INTACT Mass spectra recorded for the synthetic 120-mer Poly(A) Tail oligonucleotide: (A) MaxEnt1 charge deconvolved spectrum; (B) raw ESI-MS spectrum. The sequence of the 120-mer shown in panel A contains a cytidine incorporated at position 28, a uridine incorporated at position 56, a guanosine incorporated at position 84, and another uridine at position 112. These four residues are used for increasing the chemical stability of the Poly(A) Tail.
In the next step, the deconvolved spectra were searched automatically against a list of 20 possible modifications including the addition and subtraction of up to 10 adenosine residue masses (329.2059 Da) as shown in the INTACT Mass screenshot displayed in Figure 6. The deconvolved spectral results from the automatically generated report are shown in Figure 7. As indicated by this report, seven Poly(A) Tail species were putatively identified, with mass accuracy better than 25 ppm. In addition to the expected 120-mer Poly(A), three species containing more adenosine residues (121,122, and 123-mers), as well as three species containing fewer adenosines (117, 118, and 119-mer) were identified. The lowest identified species, a 123-mer was detected at 1.5% abundance level according to its ESI-MS response.
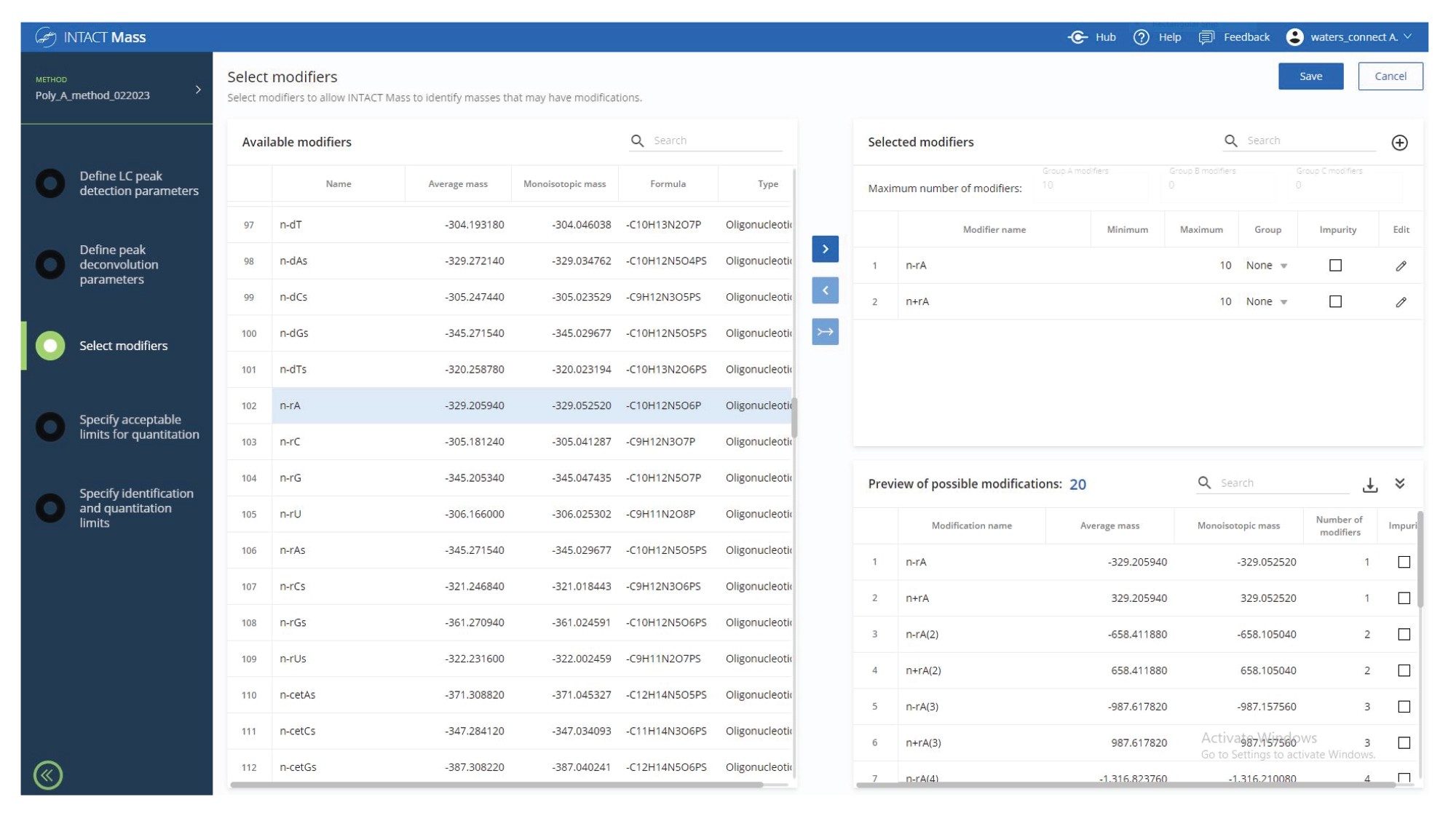 Figure 6. INTACT Mass App screenshot showing twenty possible oligonucleotide modifications, including the addition and subtraction of up to 10 adenosine masses (329.2059 Da), that could be present in the crude 120-mer synthetic Poly(A) Tail sample.
Figure 6. INTACT Mass App screenshot showing twenty possible oligonucleotide modifications, including the addition and subtraction of up to 10 adenosine masses (329.2059 Da), that could be present in the crude 120-mer synthetic Poly(A) Tail sample.
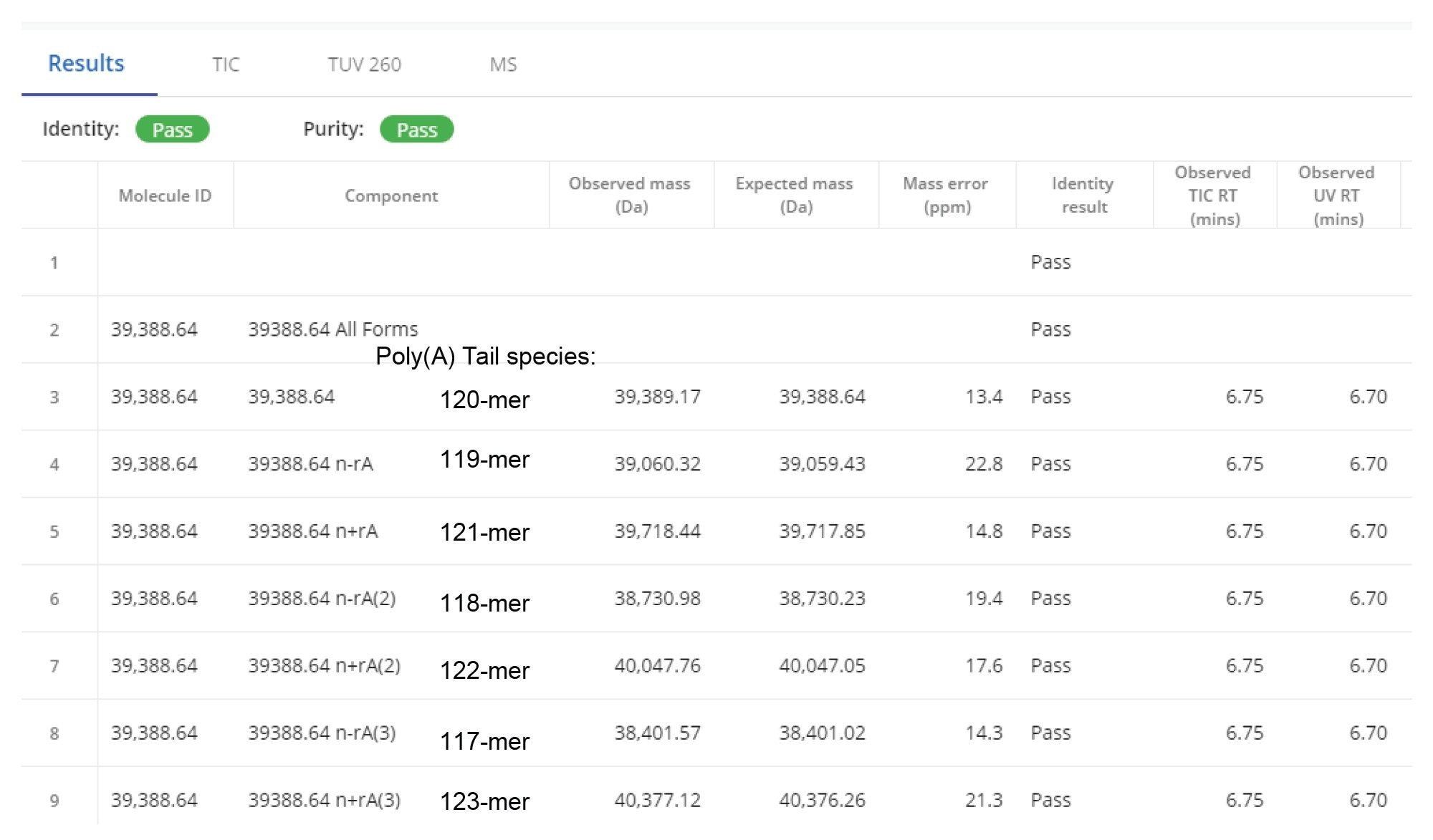 Figure 7. Section of the INTACT Mass App report displaying the processing results obtained for the analysis of the 120-mer sgRNA oligonucleotide. Seven oligonucleotide species, ranging from 117- to 123-mers were identified with mass accuracies under 25 ppm.
Figure 7. Section of the INTACT Mass App report displaying the processing results obtained for the analysis of the 120-mer sgRNA oligonucleotide. Seven oligonucleotide species, ranging from 117- to 123-mers were identified with mass accuracies under 25 ppm.
Poly(A) Tail Analysis of an mRNA Digest
The same LC-MS assay developed for the synthetic Poly(A) Tail was applied to a RNAse T1 digested Fluc (firefly luciferase) mRNA. A previous report proposed an isolation procedure of the Poly(A) Tail oligonucleotide from the complex mRNA digest mixture, using (dT)25 magnetic beads, in order to enrich this oligonucleotide fraction before subsequent LC-MS analysis.14 However, in our case, the separation of the digestion mixture, shown in Figure 8, exhibited a well resolved, abundant, Poly(A) Tail eluting as the last eluting peak in the UV and TIC chromatograms.
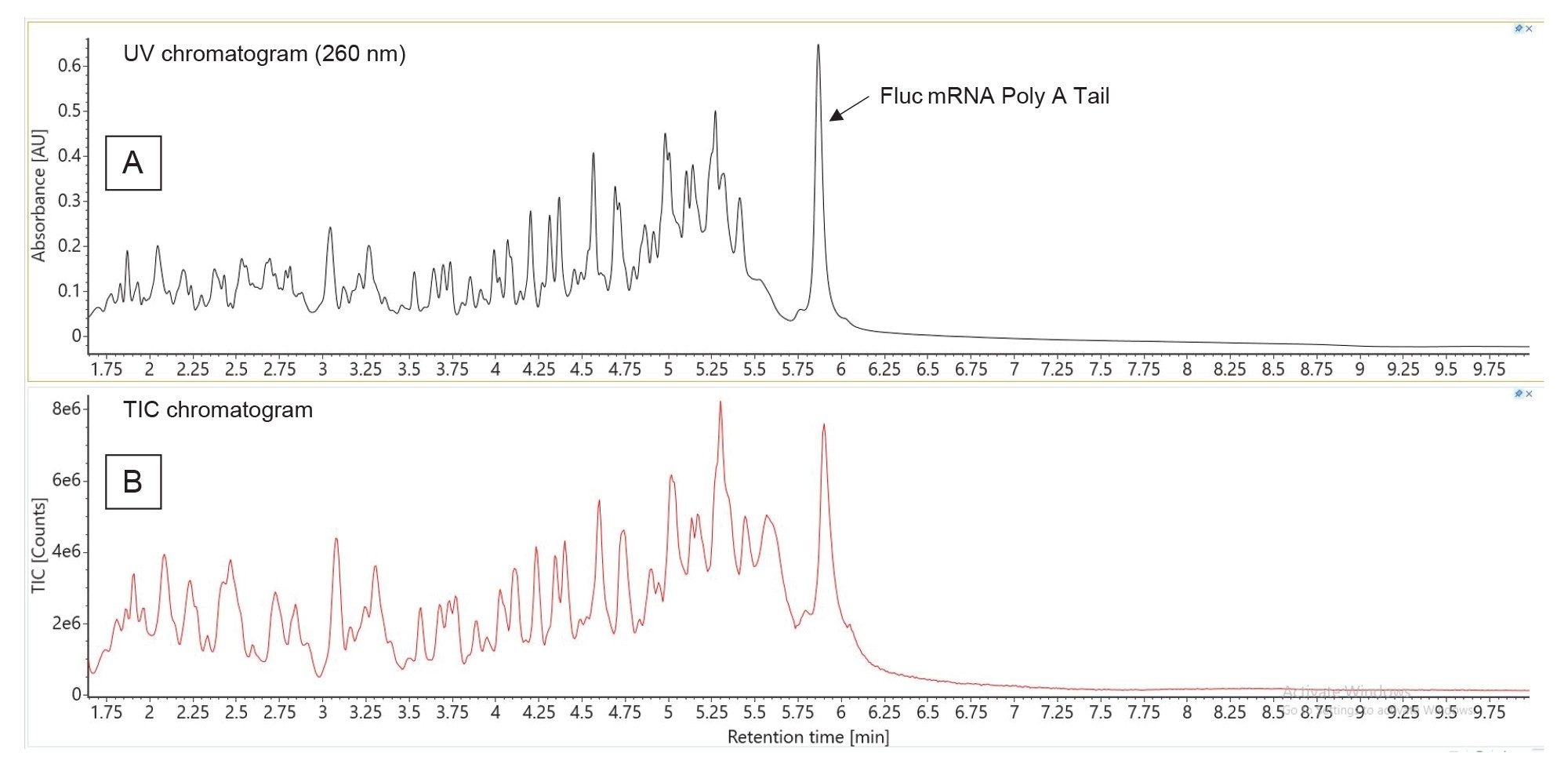 Figure 8. Separation of the RNase T1 digested Fluc mRNA: (A) UV chromatogram recorded at 260 nm; (B) TIC chromatogram recorded on the BioAccord LC-MS System.
Figure 8. Separation of the RNase T1 digested Fluc mRNA: (A) UV chromatogram recorded at 260 nm; (B) TIC chromatogram recorded on the BioAccord LC-MS System.
The corresponding deconvolved ESI-MS spectrum (Figure 9), indicates a relatively high Poly(A) heterogeneity, with oligonucleotides in the range of 122–132-mers, based on the presence of many signals spaced by one adenosine mass (329.2 Da). Following INTACT Mass processing with Custom deconvolution parameters, almost all of these eleven Poly(A) variants were measured with mass accuracies better than 30 ppm, as shown in the results displayed in Figure 10.
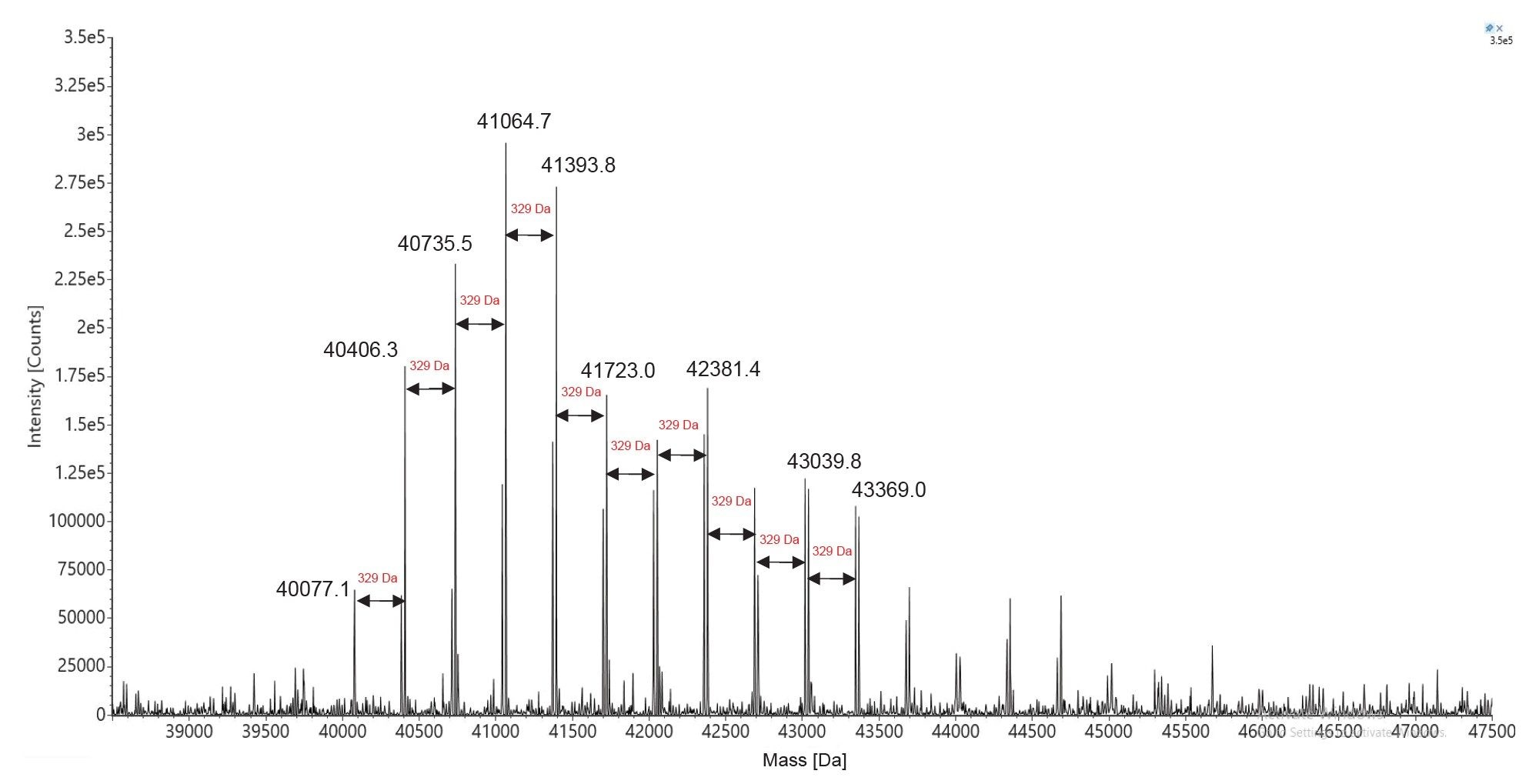 Figure 9. MaxEnt1 charge deconvolved spectrum of the Poly(A) Tail oligonucleotide mixture resulted from RNase T1 digestion of the Fluc mRNA. The Poly(A) Tail oligonucleotide has a wide dispersity, with up to eleven adenosine masses added to the first detected species, a 122-mer.
Figure 9. MaxEnt1 charge deconvolved spectrum of the Poly(A) Tail oligonucleotide mixture resulted from RNase T1 digestion of the Fluc mRNA. The Poly(A) Tail oligonucleotide has a wide dispersity, with up to eleven adenosine masses added to the first detected species, a 122-mer.
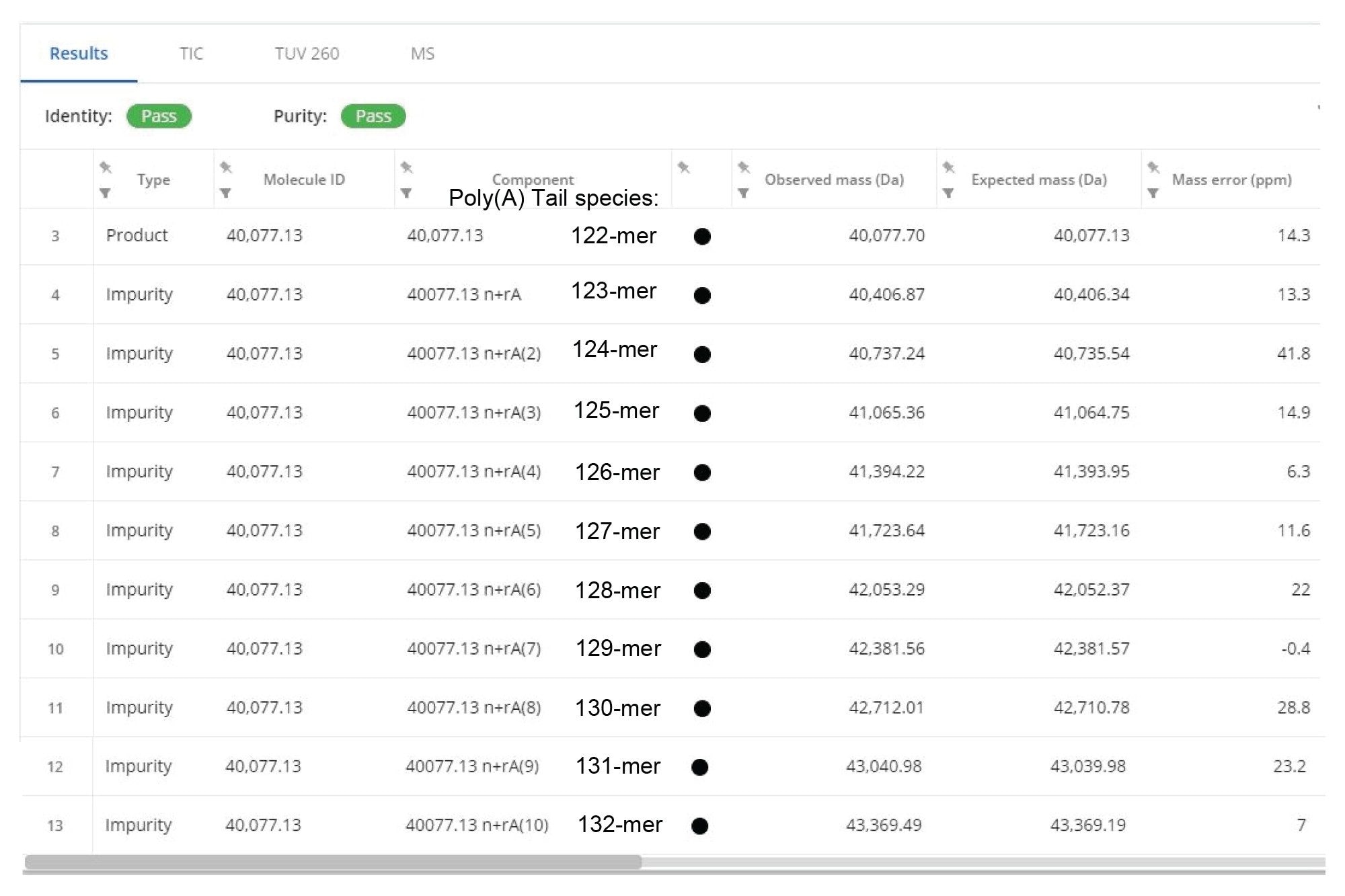 Figure 10. Section of the INTACT Mass App report displaying the processing results obtained for the analysis of the Fluc mRNA Poly(A) Tail. Eleven oligonucleotide species, ranging from 122- to 132-mers were identified with mass accuracies better than 30 ppm.
Figure 10. Section of the INTACT Mass App report displaying the processing results obtained for the analysis of the Fluc mRNA Poly(A) Tail. Eleven oligonucleotide species, ranging from 122- to 132-mers were identified with mass accuracies better than 30 ppm.
In a different graphical representation, the ESI-MS signals and their corresponding intensities were plotted in Figure 11 to calculate the average mass of the Poly(A) Tail of the Fluc mRNA which corresponds to a 126.5-mer. Both the Poly(A) dispersity as well as the average mass measurement are important quality attributes for therapeutic mRNA molecules. The average Poly(A) Tail length from the IP-RP LC-MS data presented here correlated well with the value obtained from the SEC-UV method.20 Thus, the assay developed on the synthetic Poly(A) Tail oligonucleotide was demonstrated to be effective for analysis of Poly(A) Tails enzymatically cleaved from prophylactic/therapeutic mRNAs.
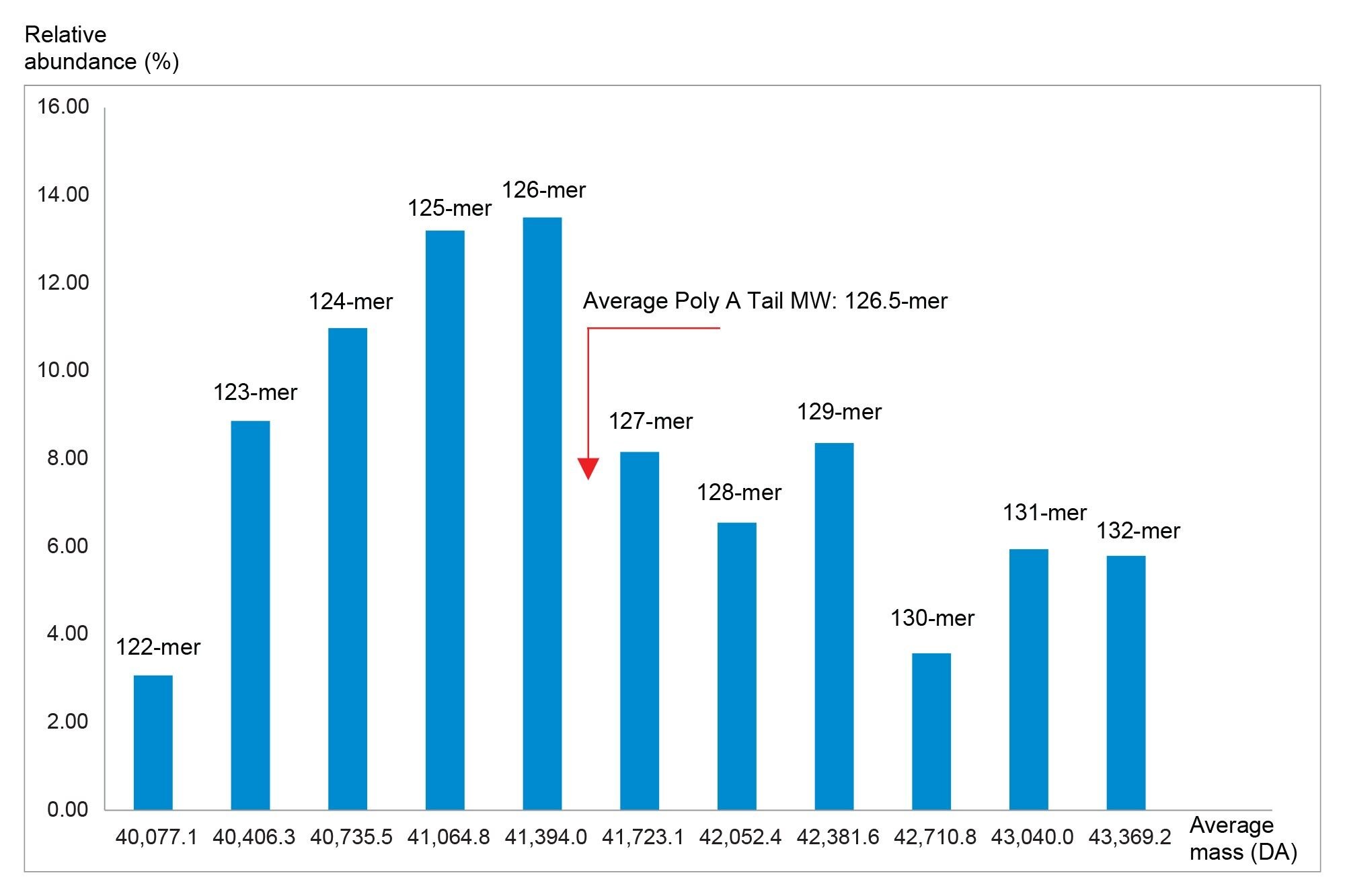 Figure 11. Distribution of the average ESI-MS spectral intensity versus the Poly(A) Tail length for the Fluc mRNA.
Figure 11. Distribution of the average ESI-MS spectral intensity versus the Poly(A) Tail length for the Fluc mRNA.
Conclusion
- An IP-RP LC-MS assay developed on the BioAccord LC-MS System provides length and heterogeneity information, as well as the average molecular weight for the Poly(A) Tail oligonucleotide region of prophylactic/therapeutic mRNAs
- Various co-eluting Poly(A) Tail oligonucleotides can be measured accurately with mass accuracies of better than 25 ppm on the bench-top BioAccord LC-MS System operated under the compliance-ready waters_connect informatics platform
- An LC-MS assay developed for a synthetic Poly(A) analogue, can be successfully applied for the analysis of real Poly(A) 3’-end fragments produced following the enzymatic digestion of prophylactic/therapeutic mRNAs mRNAs
References
- Jackson NAC, Kester KE, Casimiro D, Gurunathan S, DeRosa F. The Promise of mRNA Vaccines: A Biotech and Industrial Perspective, npj Vaccines, 2020, 11, 1–6.
- Sahin U, Kariko K, Tureci O. mRNA-Based Therapeutics – Developing a New Class of Drugs, Nat Rev, 2014, 13, 759–780.
- Sheets MD, Wickens W. Two Phases in the Addition of a Poly(A) Tail, Genes & Develop, 1989, 3, 1401–1412.
- Dreyfus M, Regnier P. The Poly(A) Tail of mRNAs: Bodyguard in Eukaryotes, Scavenger in Bacteria, Cell, 2002, 111, 611–613.
- Chang H, Lim J, Ha M, Kim VN. TAIL-seq: Genome-Wide Determination of Poly(A) Tail Length and 3′ End Modifications, Mol Cell, 2014, 53, 1044–1052.
- Nicholson AL, Pasquinelly AE. Tales of Detailed Poly(A) Tails, Trends Cell Biol, 2019, 29, 191–200.
- Jiang T, Yu N, Kim J, Murgo JR, Kissai M, Ravichandran K, Miracco EJ, Presnyak V, Hua S. Oligonucleotide Sequence Mapping of Large Therapeutic mRNAs via Parallel Ribonuclease Digestions and LC-MS/MS, Anal Chem, 2019, 91, 8500–8506.
- Vanhinsbergh C, Criscuolo A, Sutton JN, Murphy K, Williamson AJK, Cook K, Dickman MJ. Characterization and Sequence Mapping of Large RNA and mRNA Therapeutics Using Mass Spectrometry, Anal Chem, 2022, 94, 7339–7349.
- Muttach F, Muthmann N, Rentmeister A. Synthetic mRNA Capping, Beilstein J Org Chem, 2017, 13, 2819–2832.
- Beverly M, Dell A, Parmar P, Houghton L. Label-Free Analysis of mRNA Capping Efficiency Using RNse H probes and LC-MS, Anal Bioanal Chem, 2016, 408, 5021–5030.
- Nguyen J, Chan SH, Roy B, Gilar M, Robb B, Chen W, Lauber MA. Rapid Analysis of Synthetic mRNA Cap Structure Using Ion-Pairing RPLC with the BioAccord LC-MS System, Waters Application Note, 720007329, 2021.
- Muthmann N, Spacek P, Reichert D, van Dulmen M, Rentmeister A. Quantification of mRNA Cap-Modifications by Means of LC-QqQ-MS, Methods, 2022, 203, 196–206.
- Chan SH, Whipple JM, Dai N, Kelley TM, Withers K, Tzertzinis G, Correa Jr IR, Robb GB. RNase-H-based Analysis of Synthetic mRNA 5’ Cap Incorporation, RNA, 2022, 28, 1144–1155.
- Beverly M, Hagen C, Slack O. Poly A Tail Length Analysis of In Vitro Transcribed mRNA by LC-MS, Anal Bioanal Chem, 2018, 410, 1667–1677.
- Strezsak SR, Pimentel AJ, Hill IT, Beuning PJ, Skizim NJ. Novel Mobile Phase to Control Charge States and Metal Adducts in the LC/MS for mRNA Characterization Assays, ACS Omega Reviews, 2022, 7, 22181–22191.
- Doneanu C E, Fox J, Harry E, Knowles C, Yu Y Q, Fredette J, Chen W. An Automated Compliance-Ready LC-MS Workflow for Intact Mass Confirmation and Purity Analysis of Oligonucleotides, Waters Application Note, 720006820, 2020.
- Doneanu C E, Fox J, Harry E, Knowles C, Yu Y Q, Fredette J, Chen W. Intact Mass Confirmation Analysis on the BioAccord LC-MS System for a Variety of Extensively Modified Oligonucleotides, Waters Application Note, 720007028, 2020.
- Doneanu C E, Knowles C, Fox J, Harry E, Yu Y Q, Fredette J, Chen W. Analysis of Oligonucleotide Impurities on the BioAccord System with ACQUITY Premier, Waters Application Note, 720007301, 2021.
- Doneanu C E, Boyce P, Shion H, Fredette J, Berger S J, Gastall H, Yu Y Q. LC-MS Analysis of siRNA, Single Guide RNA and Impurities Using the BioAccord™ System with ACQUITY™ Premier and New Automated INTACT Mass Application, Waters Application Note, 720007546, 2022.
- Gilar M. Size-Exclusion Chromatography Method for Poly(A) Tail Analysis of mRNA, Waters Application Note, 720007853, 2023.
- Shion H, Boyce P, Berger S J, Yu Y Q. INTACT Mass™ - a Versatile waters_connect™ Application for Rapid Mass Confirmation and Purity Assessment of Biotherapeutics, Waters Application Note, 720007547, 2022.
- Ferrige AG, Seddon MJ, Green BN, Jarvis SA, Skilling J. Disentangling Electrospray Spectra With Maximum Entropy, Rapid Commun. Mass Spectrom., 1992, 6, 707–711.
720007925, August 2023