Cyclic™ Ion Mobility Unravels Sites of Chemically Induced Deamidation
這是一篇應用簡報,不含詳細的實驗內容章節。
Abstract
Acid deamidation can mimic the action of tissue transglutaminase in deamidation of specific peptide glutamine residues, and possibly increase the immunotoxicity of the deamidated gluten food ingredient and convey an increased risk to those with coeliac disease. Peptides with multiple deamidation sites may co-elute under the same chromatographic peak and therefore other methods of site determination are required such as ion mobility mass spectrometry, increasing confidence in profiling these post-translational modifications (PTMs).
Mass spectrometry profiling of peptides subject to chemical deamidation revealed a complex pattern and number of deamidation sites within the peptide sequence due to the large number of glutamine residues present. Analysis with the SELECT SERIES™ Cyclic IMS using multi-pass ion mobility, allowed for the separation of peptides with a similar number of deamidated residues, however with different sites due to the unique conformational changes.
Benefits
- The SELECT SERIES Cyclic IMS can resolve native and singly deamidated species using just a single pass of the mobility device
- The multi-pass functionality (IMSn) with increased separation length, allowing for differentiation between species consisting of differing amounts of deamidated residues
- Gas phase separation of near isobaric species in the mobility cell can facilitate deconvolution of complex chromatograms alongside complex spectra
Introduction
Immune-mediated adverse reactions to food include IgE-mediated allergy and coeliac disease, both of which can be elicited by ingestion of gluten containing products. In coeliac disease, epitopes within these proteins known as coeliac toxic motifs can reach the gut intact and specific glutamine residues are deamidated to glutamic acid by tissue transglutaminase, increasing their immunotoxicity, and resulting in a +0.984 Da mass shift.1 Deamidation can also be induced through treatment with acids and indeed, deamidated gluten is used as a food ingredient as it is more dispersible in water than the non-deamidated form.2 However, the pattern of deamidation caused by treatment with food-safe acids such as tartaric acid (TA) compared to tissue transglutaminase is unclear, and therefore any increase in immunotoxicity compared to the native protein is also unclear.
Localization of sites of deamidation within peptides consisting of multiple, susceptible residues is difficult since different species may co-elute under the same chromatographic peak, confounded by the overlapping isotopic distributions of the different deamidated species. Therefore, other methods are often required to facilitate site determination such as implementing ion mobility which has the capability to separate differing structures and potentially deconvolve the overlapping distributions and differentiate between very similar masses.
The SELECT SERIES Cyclic IMS (Figure 1) employs a closed-loop ion mobility cell allowing for multiple passes of ions and undertaking IMSn experiments, whilst also containing an orthogonal time-of-flight (ToF) analyzer capable of 100,000 FWHM resolving power facilitating the differentiation of near isobaric species such as differentially deamidated peptides.
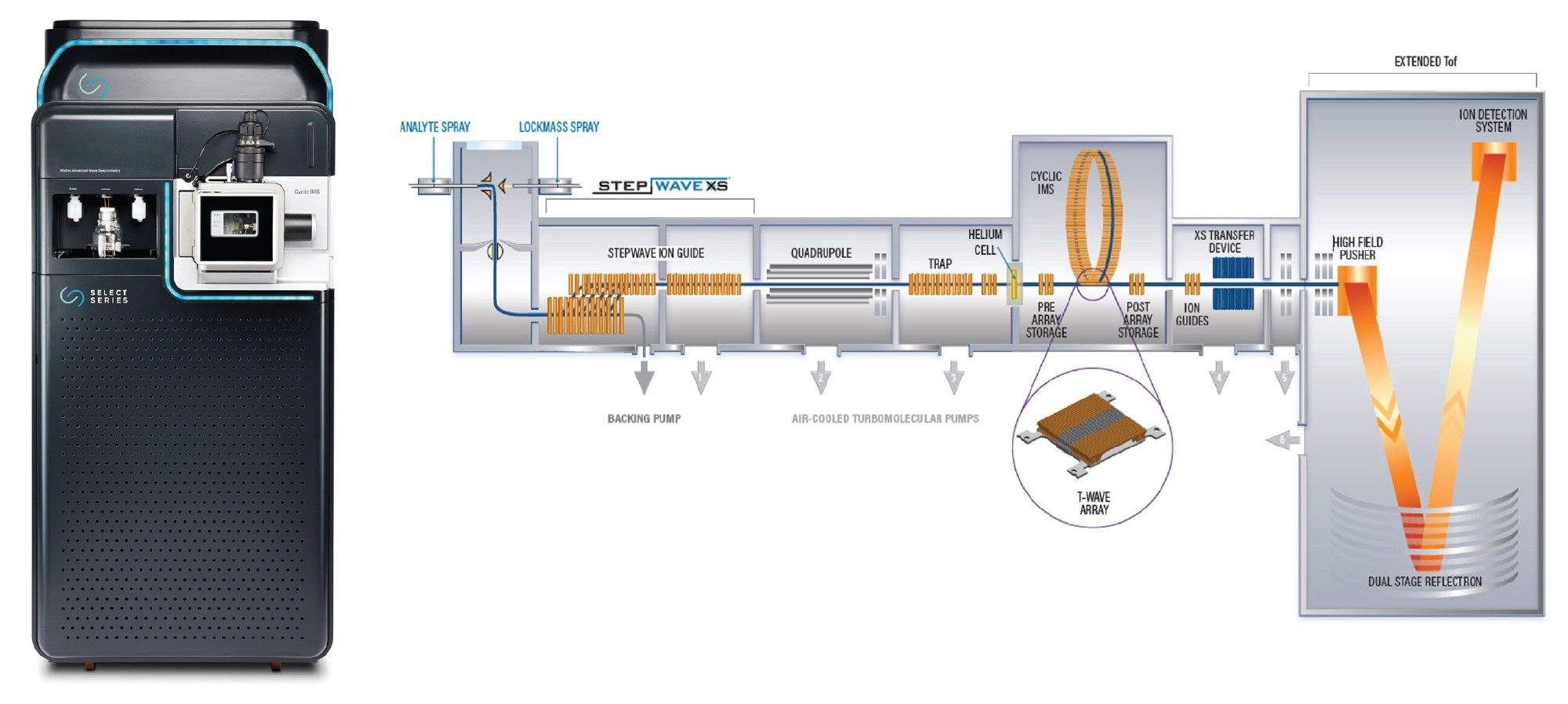 Figure 1. SELECT SERIES Cyclic IMS System (left) and a schematic of the SELECT SERIES Cyclic IMS System (right).
Figure 1. SELECT SERIES Cyclic IMS System (left) and a schematic of the SELECT SERIES Cyclic IMS System (right).
Results and Discussion
Initially, the peptide ASIVAGISQ, derived from a chymotryptic digest of wheat γ-gliadin and with only one potential site for deamidation, was chemically deamidated by incubation with tartaric acid. The resulting products were analyzed using the SELECT SERIES Cyclic IMS. Due to the small mass difference between the C13 isotope of the native peptide and C12 isotope of the deamidated product (+0.006 Da), it can be difficult to differentiate between the species using conventional MS techniques.
However, the two species could be separated by ion mobility after a single pass of the cyclic device (Figure 2A). After three passes for a drift length of ~3 m the two species were baseline separated (Figure 2A), and the extracted spectra from those separated peaks (Figure 2B) clearly demonstrate the presence of the deamidated and native product with monoisotopic m/z of 903.48 and 902.50, respectively.
Therefore, the distinction between native and singly deamidated species is clearly demonstrated by cyclic ion mobility using a drift length of ~1 m only and without the need for further characterization.
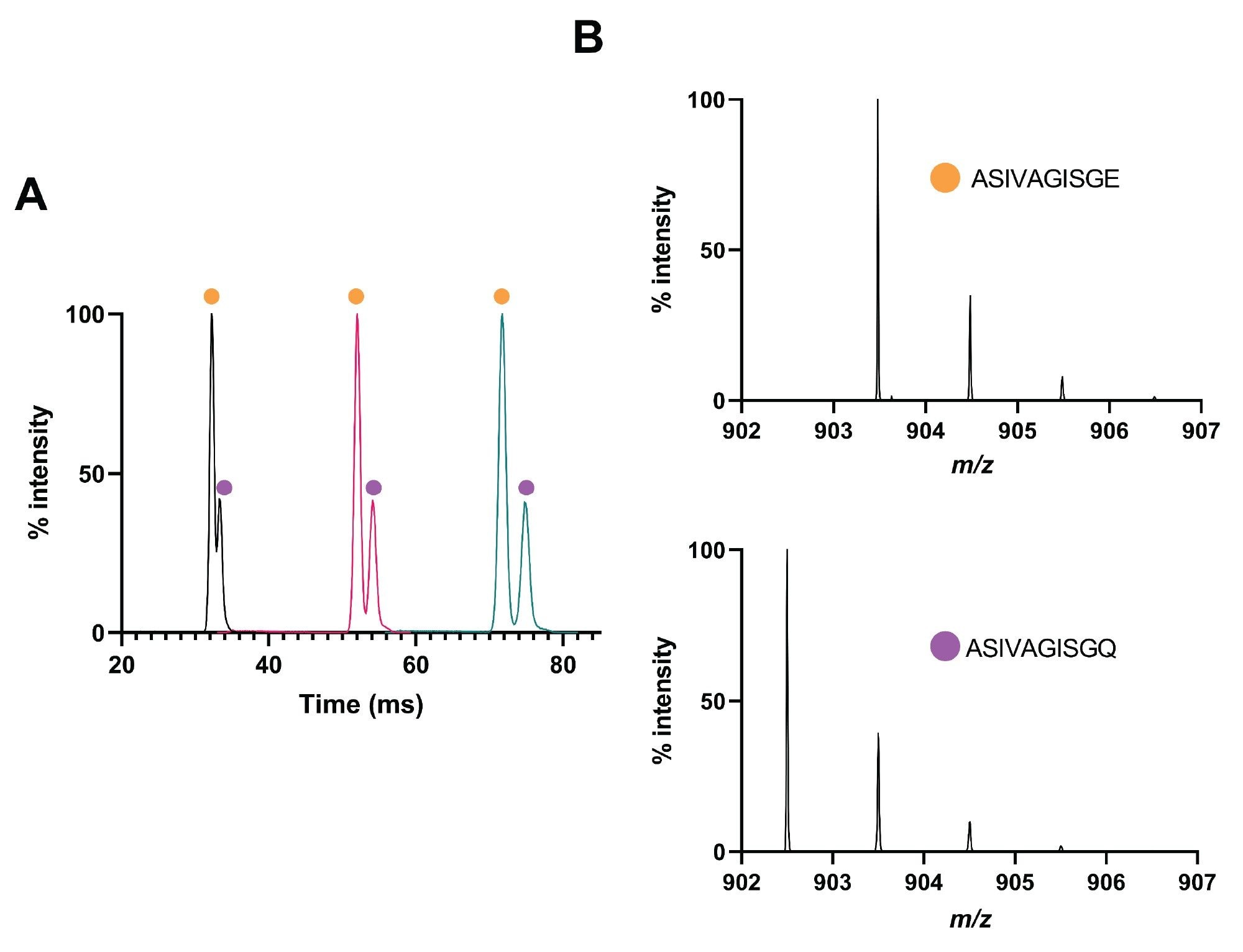 Figure 2. Analysis of peptide ASIVAGISQ and deamidated product. (A) Mobilogram demonstrating separation of native (purple circle) and deamidated peptide (orange circle) after one, two, and three passes of the IMS cell. (B) Mass spectra associated to each peak from (A) demonstrating the mass associated with the native or deamidated product.
Figure 2. Analysis of peptide ASIVAGISQ and deamidated product. (A) Mobilogram demonstrating separation of native (purple circle) and deamidated peptide (orange circle) after one, two, and three passes of the IMS cell. (B) Mass spectra associated to each peak from (A) demonstrating the mass associated with the native or deamidated product.
The peptide GIIQPQQPAQL was then subject to chemical deamidation and analysis using the SELECT SERIES Cyclic IMS. Due to the presence of four glutamine residues, there were 16 possible species (one native, four with one deamidation, six with two deamidations, four with three deamidations, and one with four deamidations). Such complexity necessitated multiple passes of the ion mobility cell, and indeed, three passes were required before two peaks were distinguishable, and five passes to further separate them (Figure 3A). Extracted masses from the separated arrival time distributions, demonstrated a peak at ~101 ms containing a mix of species primarily with at least one deamidation, and a second peak at ~98 ms containing primarily species with at least two deamidations (Figure 3B). A peak associated with the monoisotopic species of the native peptide (596.83 m/z) was observed although at a very low intensity, suggesting the majority of the peptide underwent at least partial conversion to the deamidated form.
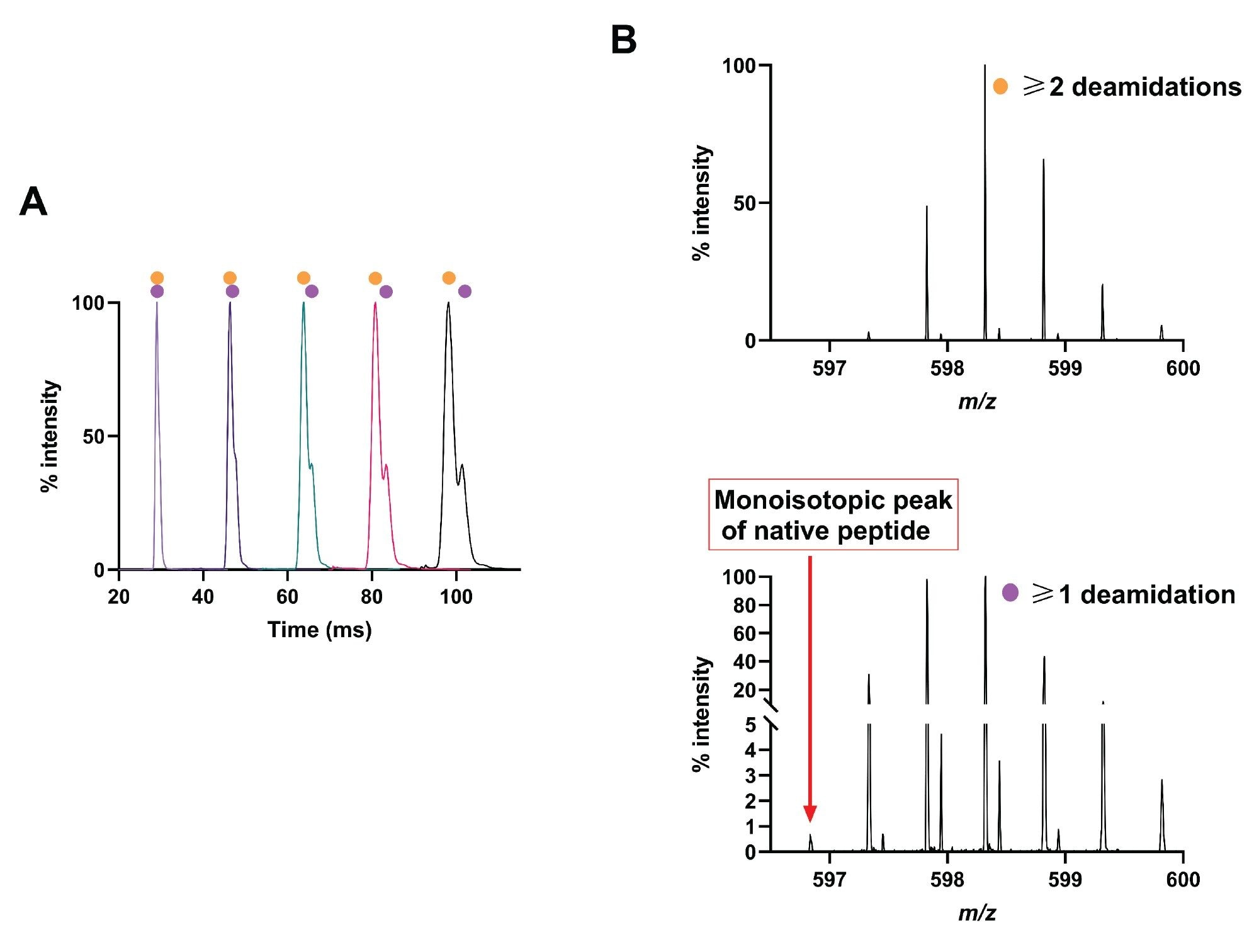 Figure 3. Analysis of peptide GIIQPQQPAQL and deamidated products. (A) Mobilogram demonstrating separation of species with ≥1 (purple circle) and ≥2 deamidations (orange circle) after one, two, three, four, and five passes of the IMS cell. (B) Mass spectra associated to each peak from (A) demonstrating the mass associated with the different species.
Figure 3. Analysis of peptide GIIQPQQPAQL and deamidated products. (A) Mobilogram demonstrating separation of species with ≥1 (purple circle) and ≥2 deamidations (orange circle) after one, two, three, four, and five passes of the IMS cell. (B) Mass spectra associated to each peak from (A) demonstrating the mass associated with the different species.
Focus was then applied on species with only a single deamidated glutamine residue and monoisotopic m/z of 597.34 which would also include the first isotope of the native peptide. Four distinct peaks emerged after five passes in the ion mobility cell (Figure 4).
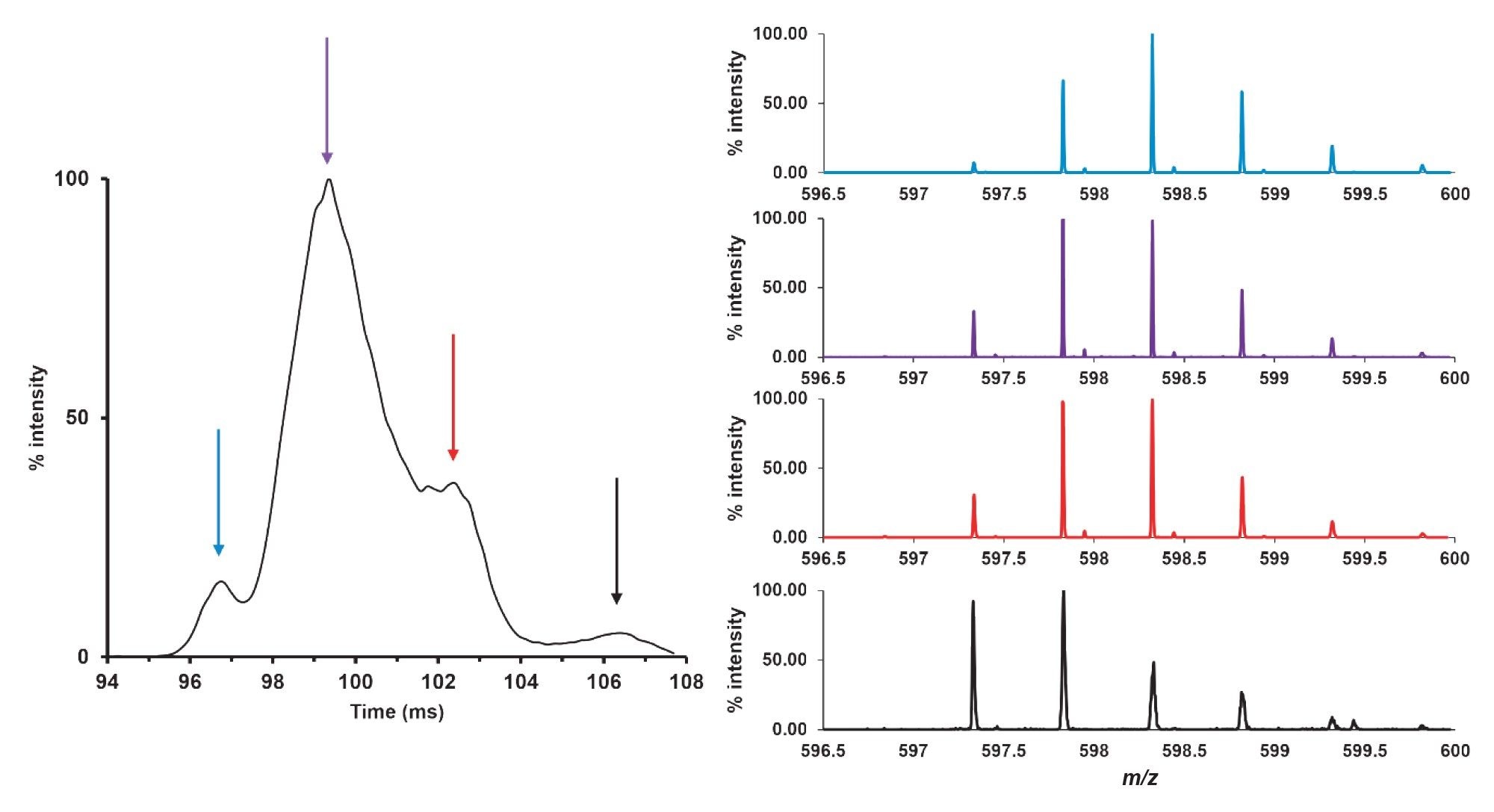 Figure 4. Extracted ion mobilogram at 597.34 m/z corresponding to the peptide GIIQPQQPAQL with one deamidated glutamine residue (out of a possible four). Based on drift time after five passes, four different species could be separated.
Figure 4. Extracted ion mobilogram at 597.34 m/z corresponding to the peptide GIIQPQQPAQL with one deamidated glutamine residue (out of a possible four). Based on drift time after five passes, four different species could be separated.
The mobility peak designated by the blue arrow and the extracted mass spectrum (also in blue) demonstrate a population largely consisting of species with two or more deamidations. Peaks and spectrum represented by purple and red with increasing arrival time show a shift towards species with a decreasing number of deamidations as evidenced by the increase in intensity at 597.34 m/z (one deamidation) and decrease in relative intensity of the peak at 598.3 m/z (three deamidations). The trend continued as the arrival time increased, with the mobility peak indicated by the black arrow containing a larger proportion of species with one and two deamidations. Therefore, as more of the glutamine residues within the sequence were deamidated to glutamic acid, the arrival time decreased contrary to the normal pattern of increased arrival time with increased m/z.
The applicability of ion mobility and necessity of the ability to increase drift time to characterize the extent of deamidation in peptides with multiple susceptible residues is therefore demonstrated.
Conclusion
The SELECT SERIES Cyclic IMS could differentiate between a native peptide and one with a single deamidation in a single pass of the IMS cell. Multi-pass capabilities to increase the drift length and perform IMSn experiments using the Cyclic IMS cell allowed for differentiation between sites of deamidation, where multiple susceptible residues existed.
The applicability of ion mobility and particularly the ability of the Cyclic IMS to increase drift time to characterize the degree of deamidation in peptides with multiple susceptible residues is shown.
Exploiting differences in the structure of PTMs using ion mobility adds another dimension of confidence to the characterization of such modifications. The benefits of this technique could also contribute to profiling other PTMs such as phosphorylation, making ion mobility a key analysis tool in describing protein biomarkers related to human disease and their activation through phosphorylation.
References
- Sollid LM et al. Update 2020: nomenclature and listing of celiac disease relevant gluten epitopes recognized by CD4+ T cells. Immunogenetics. 2020. 10.1007/s00251-019-01141-w.
- Yongquan W et al. Improving solubility and emulsifying property of wheat gluten by deamidation with four different acids: Effect of replacement of folded conformation by extended structure. Food Hydrocolloids. 2017. 10.1016/j.foodhyd.2017.04.013.
Featured Products
720008215, January 2024