Cursory Analysis of Nucleic Acid Components in Adeno-Associated Virus (AAV) Samples Using Anion-Exchange Chromatography (AEX)
Abstract
It is important to analyze adeno-associated virus (AAV) therapeutics for contaminating nucleic acid impurities because of their implications on efficacy and safety. The integrity of an AAV genome should be checked for nicks and truncations. Moreover, AAV samples should be monitored for leaked genomes and host cell nucleic acid impurities. In this application note, we show a unique method for analyzing AAV samples for the presence of nucleic acid components, and we use a proteinase K digestion of the AAV capsid proteins to help elucidate the origin of the species observed in native, unadulterated samples. An overlap of peaks between the native and proteinase K digested samples corroborates the nature of these species as potentially being free genomic material. With these preliminary results, we show there is promise in establishing an AEX analysis with the Protein-Pak™ Hi-Res Q Column to test an AAV sample for unencapsulated nucleic acid components and potentially even a genome integrity check.
Benefits
- Waters™ Protein-Pak Hi Res Q Column can be used to detect AAV species and nucleic acid impurities
- Proteinase K-digested of AAV samples can be used to release ssDNA genome material for native sample peak identification and for further characterization of AAV genome integrity
Introduction
Recombinant AAV has become one of the most relied upon viral vectors for gene therapy. During the genome-packaging process, contaminating genomes can be erroneously packaged into AAV capsids.1 It is, therefore, desirable to analyze the AAV genome integrity to ensure safety and efficacy of AAV products.
Genome size has conventionally been determined by agarose-gel electrophoresis and Southern blots.1 While cost-effective, these methods provide limited accuracy and precision.
Anion-exchange chromatography (AEX) separates molecules based on their differences in negative surface charges. This analytical technique is robust, reproducible, and quantitative. It requires only small amounts of sample and is easy to automate. Additionally, it allows for the isolation of fractions for further analysis. AEX has been utilized in multiple areas related to gene therapy, including AAV empty and full capsid separation, plasmid isoform separation, dsDNA fragment separation and single-guide RNA size and purity estimation.2–5 Since the single-stranded DNA (ssDNA) encapsulated inside the AAV capsid protein exhibits a strongly negative net charge, we investigated AEX for its inherent ability to separate capsids, free genome material, and potentially even other types of nucleic acid components.
In this application note, we show a unique AEX method for analyzing the genome material and nucleic acid components of an AAV sample. Both native and proteinase K digested samples were analyzed, and overlapping, strongly retained peaks were found to exist in both. This observation suggests that the species observed in the native sample may indeed be unencapsulated or leaked genomic material. With these preliminary results, we show there is promise in establishing an AEX analysis with the Protein-Pak Hi-Res Q Column to test an AAV sample for unencapsulated nucleic acid components and for investigating genome integrity.
Experimental
Sample Description
AAV9-Camklla-GFP was purchased from Vigene Biosciences with a titer of 1.61 x 10^14 GC/mL. Proteinase K was purchased from New England Biolabs (P8107S).
Sample Preparation
4 µL AAV9 capsid was digested with 1 µL proteinase K for two hours at 56 °C. The reaction mix was injected onto the Protein-Pak Hi Res Q Column for separation. This protocol should only be considered as a starting point for future investigations. Additional optimization should be considered to confirm the robustness of the digestion.
LC Conditions
|
LC system: |
ACQUITY UPLC H-Class Bio |
|
Detection: |
ACQUITY UPLC TUV Detector with 5 mm titanium flow cell |
|
wavelength: |
260 nm and 280 nm |
|
Vials: |
Polypropylene 12 x 32 mm Screw Neck Vial, with Cap and Pre-slit PTFE/Silicone Septum, 300 µL Volume, 100/pk (p/n: 186002639) |
|
Column temperature: |
30 ˚C |
|
Sample temperature: |
10 ˚C |
|
Injection volume: |
3 µL for initial analysis; 40 µL for fraction collection |
|
Flow rate: |
0.4 mL/min |
|
Mobile phase A: |
100 mM Tris-HCl |
|
Mobile phase B: |
100 mM Tris base |
|
Mobile phase C: |
3 M tetramethylammonium chloride (TMAC) |
|
Mobile phase D: |
Water |
|
Buffer conc. to deliver: |
20 mM |
Gradient Table
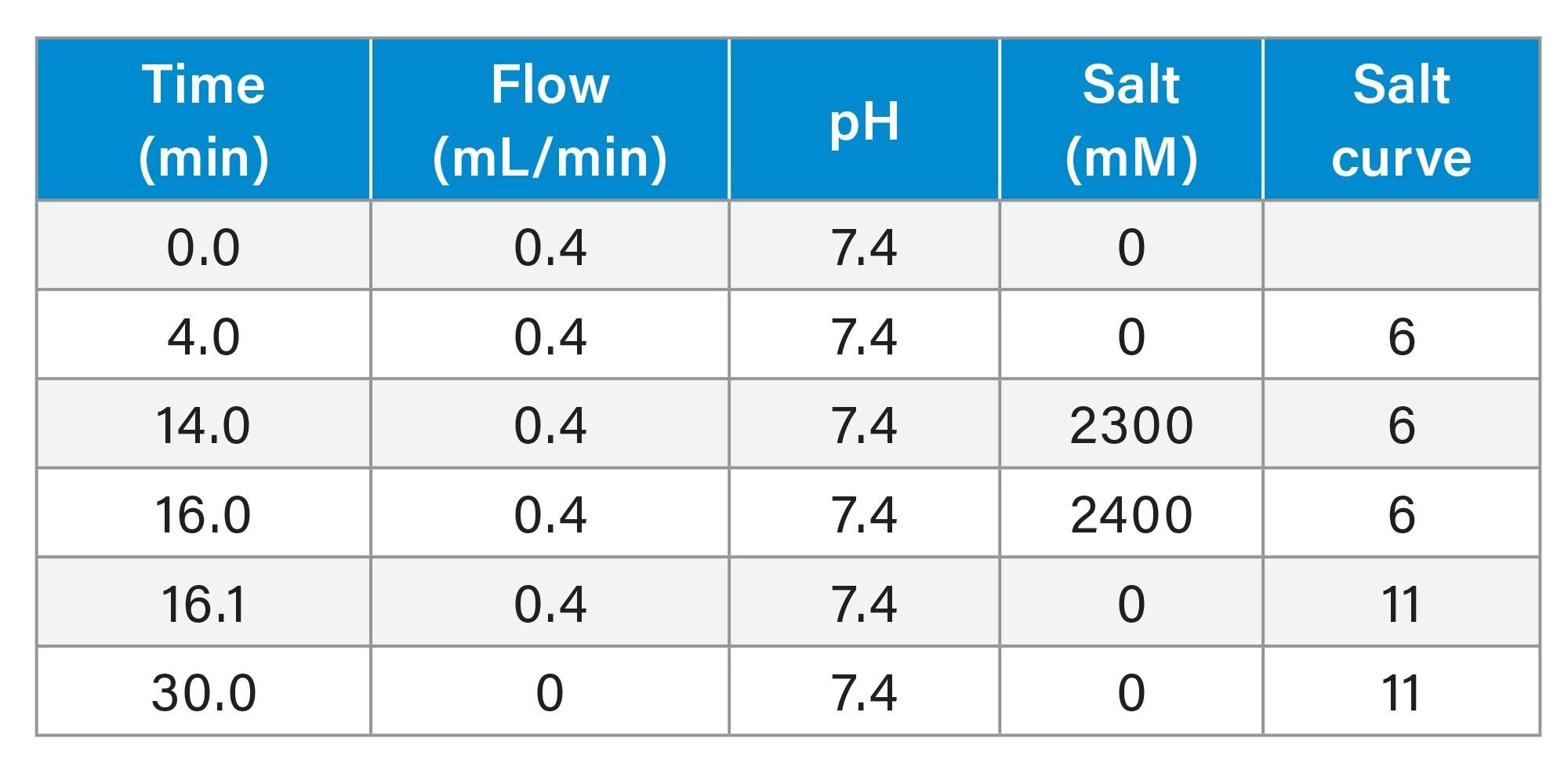 (Gradient Table) an AutoBlend Plus™ Method, Henderson-Hasselbalch derived.
(Gradient Table) an AutoBlend Plus™ Method, Henderson-Hasselbalch derived.
In the above gradient table, the buffer is 20 mM Tris pH 7.4. The initial salt concentration is set to 0 mM to ensure all the analytes are strongly bound onto the column. After four minutes of equilibration, the salt concentration increases linearly to 2300 mM in 10 minutes for a released genome analysis.
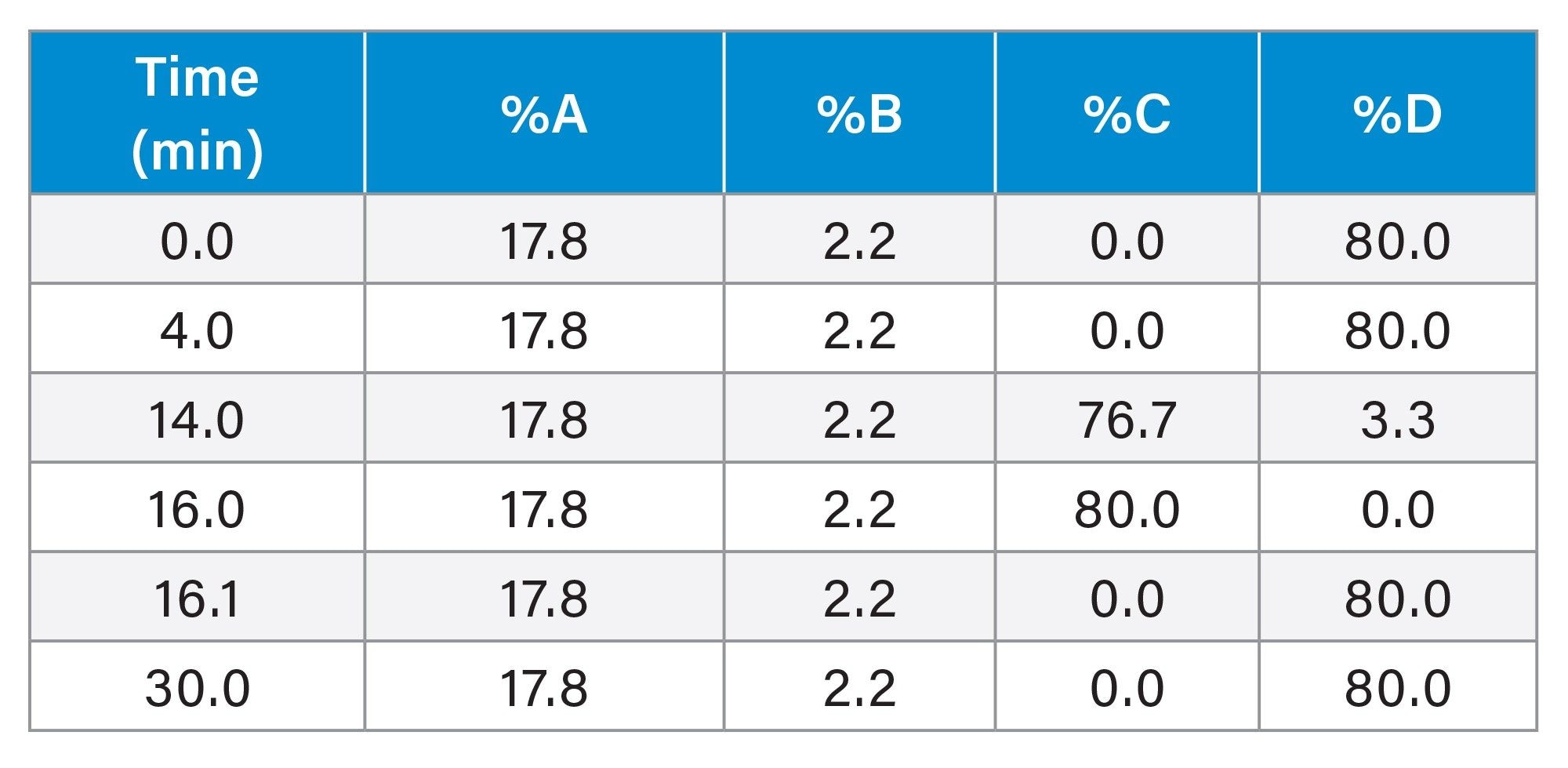
An equivalent gradient table for a generic quaternary LC system is shown below:
Data Management
|
Chromatography software: |
Empower™ 3 (FR 4) |
Results and Discussion
The ssDNA genome inside an AAV9 capsid was released by digesting the capsid protein with Proteinase K. This method is adopted from procedures used for genome titer determination.6,7 As evidenced on the anion exchange separation (Figure 1), after Proteinase K digestion, the peak area in the region where the intact capsid elutes decreased significantly, while two major peaks appeared at later retention times. Two wavelengths were used for UV detection: 260 and 280 nm. The ratio of A260/A280 is used to identify the nature of the peaks. For example, before Proteinase K digestion, A260/A280 ratio is ~1.3. This is a result of the combined absorptivity of the capsid protein and ssDNA. Since the ssDNA has higher absorbance at 260 nm and the capsid protein absorbs more strongly at 280 nm, the overall ratio of A260/A280 results ends up being in the 1.2–1.3 range. After Proteinase K digestion, the A260/A280 ratio in the capsid region (retention time between six and eight minutes) decreased to 0.9. This shows that the absorbance at 280 nm is slightly higher than the absorbance at 260 nm, which indicates that the capsid protein or peptide begins to dominate the peak and that the genomic material that used to be inside the capsid is now being retained and eluted elsewhere in the chromatogram. On the other hand, for the peaks observed in a later eluting region (retention time ~13–15 minutes), A260/A280 ratio is ~1.8, indicating that the species within are relatively pure in terms of being comprised of nucleic acid.
Additional experiments, including Charge Detection MS (CDMS) analysis on AEX fractions further confirmed that the peaks at retention times between 13 to 15 minutes are likely to be the ssDNA released from AAV capsids, including ssDNA that was likely to have been partially annealed during sample handling (data not shown). Moreover, when longer proteinase K digestions were performed, the peak area of the capsid protein elution window continued to decrease while the free DNA region continued to increase. Digestion of empty AAV capsids did not result in a peak area increase in the DNA region. Finally, fluorescence detection showed very little signal in the DNA region, consistent with the fact that DNA does not exhibit intrinsic fluorescence.
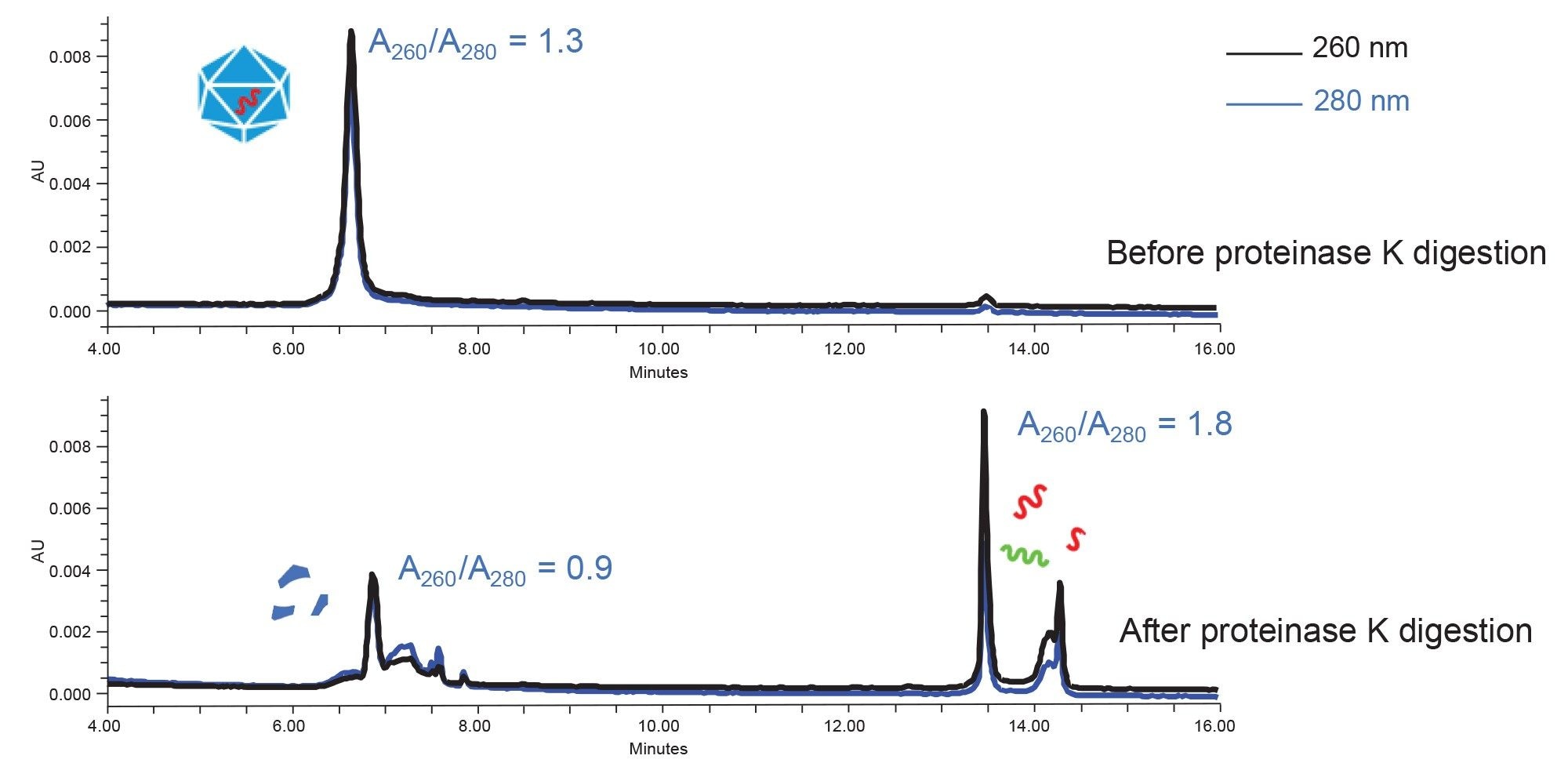 Figure 1. AEX separation of AAV9 capsid before and after proteinase K digestion. Two UV wavelengths were used for detection: 260 nm (black) and 280 nm (blue).
Figure 1. AEX separation of AAV9 capsid before and after proteinase K digestion. Two UV wavelengths were used for detection: 260 nm (black) and 280 nm (blue).
Conclusion
Anion-exchange chromatography is robust, reproducible, and quantitative, and it only requires a small amount of sample. Since AEX has proven useful for empty/full assays of intact AAVs, we thought it would also be worthwhile to test its ability to analyze AAV genomic material and map out the retention windows in which each of these components would elute. In turn, we have demonstrated that the ssDNA encapsulated within the AAV capsid can be released using proteinase K digestion and that the genome material can be well separated from capsids and capsid proteins using the Waters Protein-Pak Hi Res Q Column. It is also possible to use this column to make observations on free, unencapsulated nucleic acid components present in native AAV samples. Though this is a preliminary investigation, we show there is promise in establishing an AEX analysis with the Protein-Pak Hi-Res Q Column to test AAVs for encapsulated nucleic acid components and genome integrity upon proteinase K digestion.
References
- Hajba L and Guttman A. Recent Advances in the Analysis of Full/Empty Capsid Ratio and Genome Integrity of Adeno-associated Virus (AAV) Gene Delivery Vectors. Current Molecular Medicine 2020; 20: 1–8.
- Yang H, Koza S and Chen W. Anion-Exchange Chromatography for Determining Empty and Full Capsid Contents in Adeno-Associated Virus. Waters Application Note. 2021; 720006825.
- Yang H, Koza S and Chen W. Plasmid Isoform Separation and Quantification by Anion-Exchange Chromatography (AEX).Waters Application Note. 2021; 720007207.
- Yang H, Koza S and Chen W. Separation and Size Assessment of dsDNA Fragments by Anion-Exchange Chromatography (AEX). Waters Application Note. 2021; 720007321.
- Yang H, Koza S and Yu Y Q. Size and Purity Assessment of Single-Guide RNAs by Anion-Exchange Chromatography (AEX). Waters Application Note. 2021; 720007428.
- Werling N J, Satkunanathan S, Thorpe R, and Zhao Y. Systematic Comparison and Validation of Quantitative Real-Time PCR Methods for the Quantitation of Adeno-Associated Viral Products. HUMAN GENE THERAPY METHODS 2015; 26:82–92.
- Ai J, Ibraheim R, Tai P W L, and Gao G. A Scalable and Accurate Method for Quantifying Vector Genomes of Recombinant Adeno-Associated Viruses in Crude Lysate. HUMAN GENE THERAPY METHODS 2017; 28 (3): 139–147.
720007907, June 2023