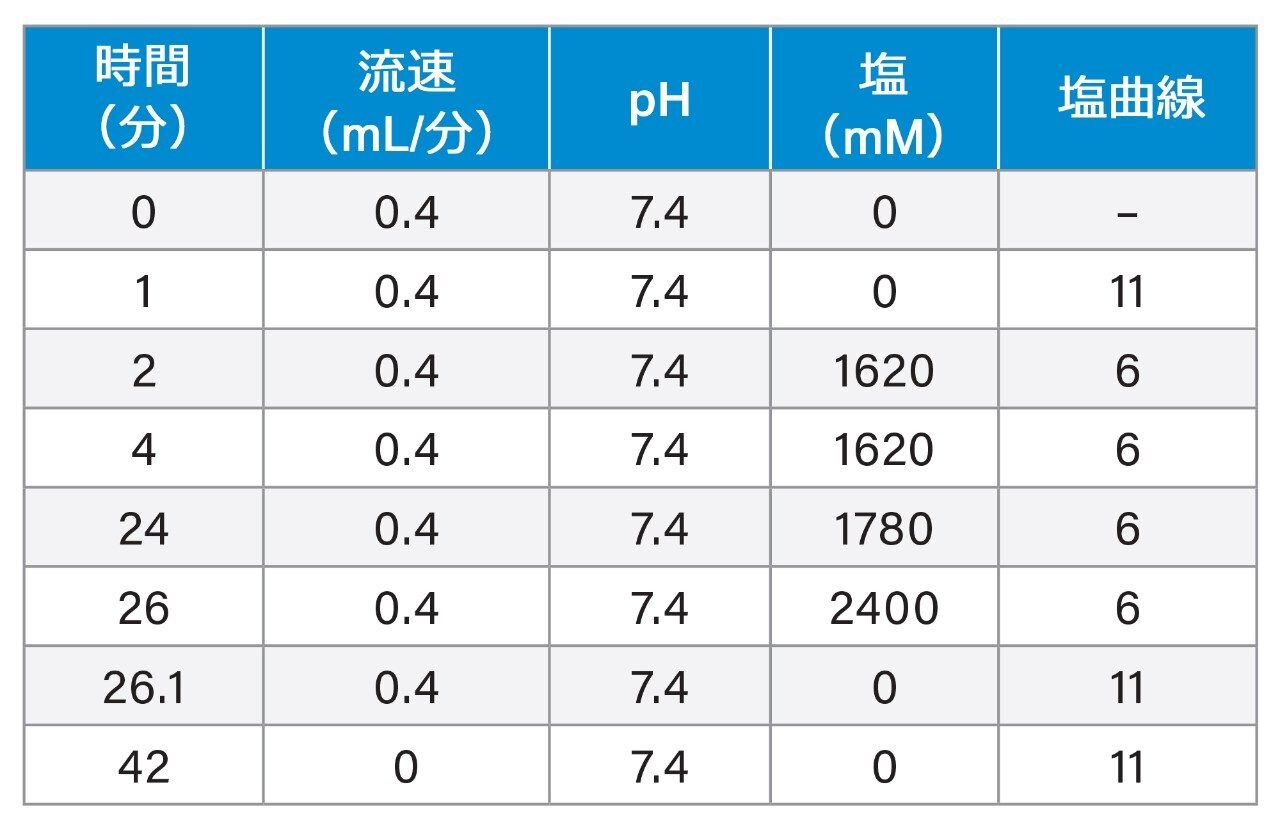
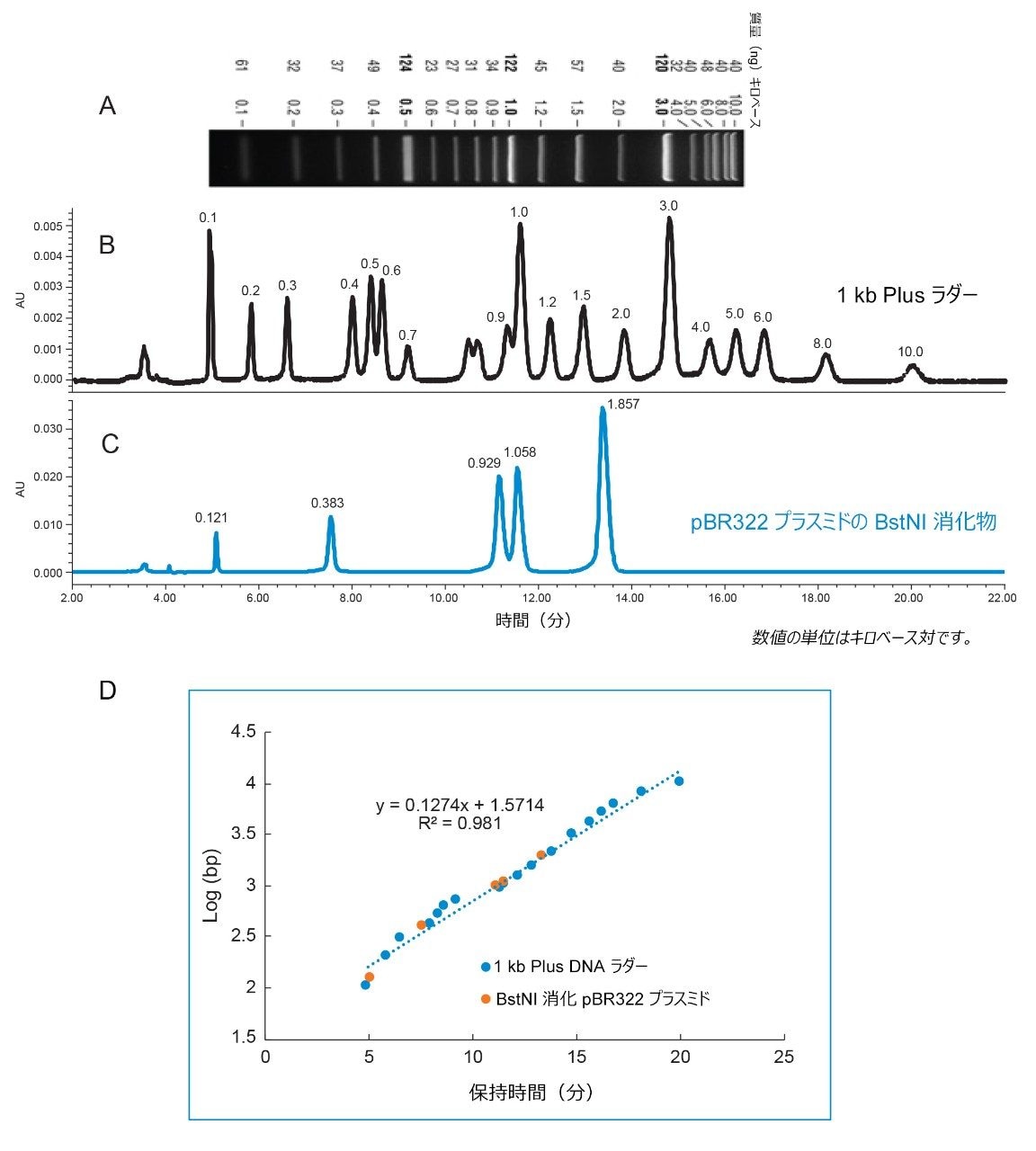
図 1B に示されているように、0.1 kbp ~ 10 kbp の範囲の一連の dsDNA フラグメントで構成される 1 kb Plus DNA ラダーが、Waters Protein-Pak Hi Res Q 強陰イオン交換カラムで分離されました。幅広いサイズのさまざまな dsDNA を分離する能力は、制限酵素消化物の分析に不可欠です。
このクロマトグラフィー分離は、New England Biolabs のウェブサイト(図 1A および 1B)に掲載されているアガロースゲル分離と一致しています。フラグメントのサイズを簡単に特定できるように、1 kb Plus dsDNA ラダー中の 1 kbp および 3 kbp のフラグメントの量を他のフラグメントの量よりも意図的に多くしました。これは、ゲル上で 1 kbp と 3 kbp のバンドが濃く明るいことから、観察できます。これに一致して、陰イオン交換クロマトグラムでの 1 kbp フラグメントと 3 kbp フラグメントのピーク面積も、他のフラグメントのピーク面積より大きくなっています。
負に帯電しているホスホジエステル基の数はヌクレオチドの数に比例するため、DNA フラグメントはそのサイズに応じて溶出するはずです。一般に、これは DNA フラグメントが短い場合(100 bp 未満)に当てはまります。DNA のサイズが大きくなると、DNA の組成が AEX 分離に影響を与える可能性があります。たとえば、A-T 含量が高いフラグメントは、鎖の長さに基づく予想時間よりも後に溶出することが示されています2,3。一部のケースでは、A-T 含量が高い短いフラグメントは、A-T 含量が低い長いフラグメントよりも後に溶出しました4。DNA は長さの順に溶出しないため、これによってサイズ評価で問題になる場合があります。これらの試みでは、AEX 分離での溶出用の塩として NaCl を使用しました。興味深いことに、NaCl 中では、A-T が多く含まれている DNA の方が、G-C が多く含まれている DNA よりも融解温度が高いことが示されています。この実験を塩化テトラメチルアンモニウム(TMAC)中で行うと、A-T が多く含まれている DNA と G-C が多く含まれている DNA の間の融解温度の差は解消されます。仮説によれば、TMAC が DNA 構造の溝に適合し、A-T 塩基対に選択的に結合して、融解温度は DNA の組成に依存しなくなります5,6。
当社での以前の研究では、核酸が含まれている分析物の溶出塩として、TMAC の方が NaCl よりも優れた分離が得られました7,8 。今回の試験では、さまざまな長さの dsDNA フラグメントを分離するための塩として、TMAC を再び実験します。多角度光散乱(MALS)で得られた予備的データでは、クロマトグラムのピークの分子量が保持時間とともに増加することが示唆されています(データは示されていません)。TMAC によって、フラグメント組成とは無関係に、サイズに基づいて DNA フラグメントを分離できる理由は、完全には明らかではありませんが、最終的な AEX 分離の結果に対する dsDNA フラグメントの組成の影響を解消する重要な役割を、TMAC が果たすと推測されます。
従来、DNA フラグメントサイズの評価は、アガロースゲルでフラグメント DNA を泳動し、バンドの位置を、同じゲルで同じ条件下でレファレンス物質として機能する DNA ラダーの位置と比較することで行います。同様に、今回の実験では、AEX メソッドでは、1 kb Plus DNA ラダーと一緒に同じクロマトグラフィー条件で分析することによって、保持時間を用いて DNA フラグメントのサイズを評価できることを実証します(図 1B と 1C)。図 1D に、dsDNA フラグメントの log(bp) 対保持時間のプロットが示されています。青色の点は 1 kb Plus DNA ラダーのデータポイントで、オレンジ色の点は BstNI 消化 pBR322 プラスミドのデータポイントです。1 kb Plus DNA ラダーによる線形回帰では、サイズの対数と保持時間の間に強い相関関係が示されています(R2 = 0.981)。このプロットを使用して、制限酵素フラグメントのサイズが保持時間から計算されます。 パーセント誤差(%)は、式 {(計算サイズ - 理論サイズ)/理論サイズ} を用いて計算されます。pBR322 プラスミド由来のほとんどの制限酵素フラグメントの測定は、誤差が 11% 未満でした。このことは、プロットのトレンドライン上または非常に近接しているオレンジ色の点によって強調されています。最初の制限酵素フラグメント(0.121 kbp)の % 誤差は、他の DNA フラグメントの % 誤差よりも大きくなっています。この偏差は、1 kb Plus DNA ラダー(0.1 kbp)で早く溶出するフラグメントのデータポイント(トレンドラインからも遠く離れている)に起因する可能性があります。