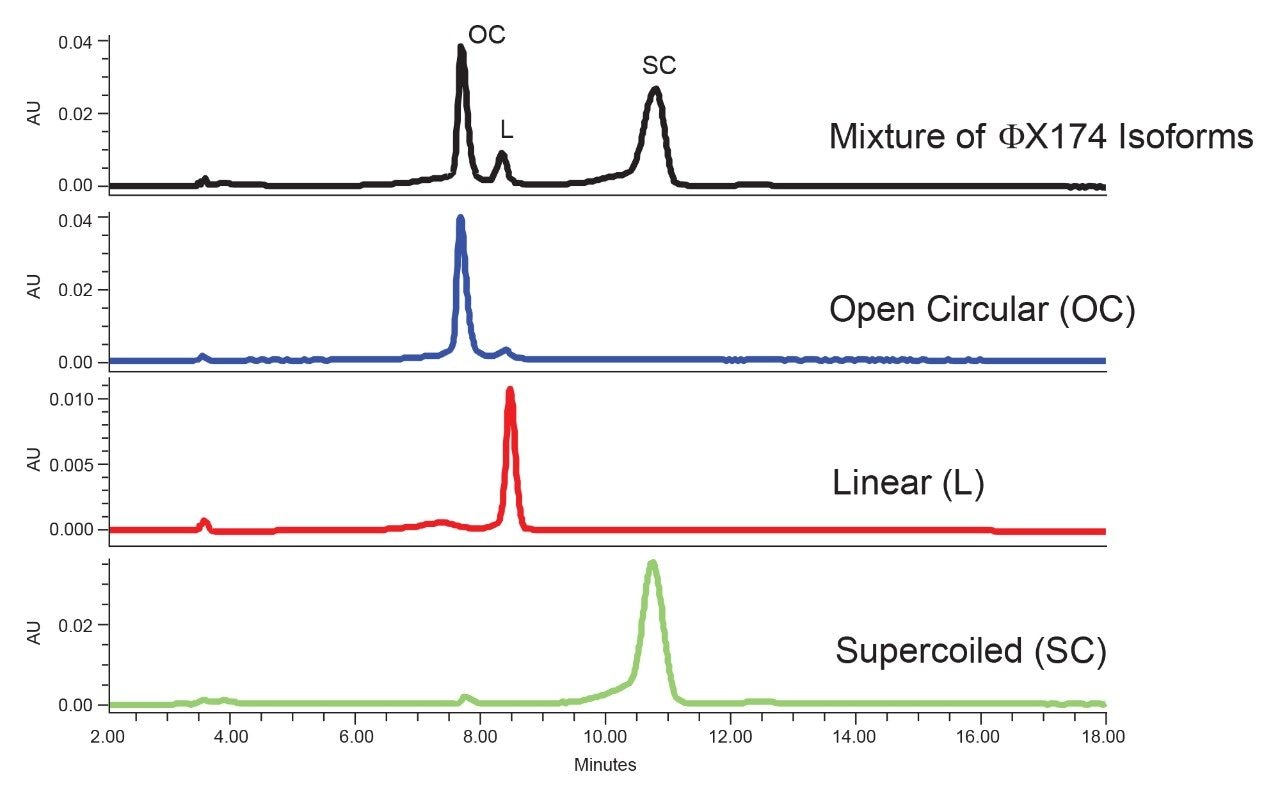
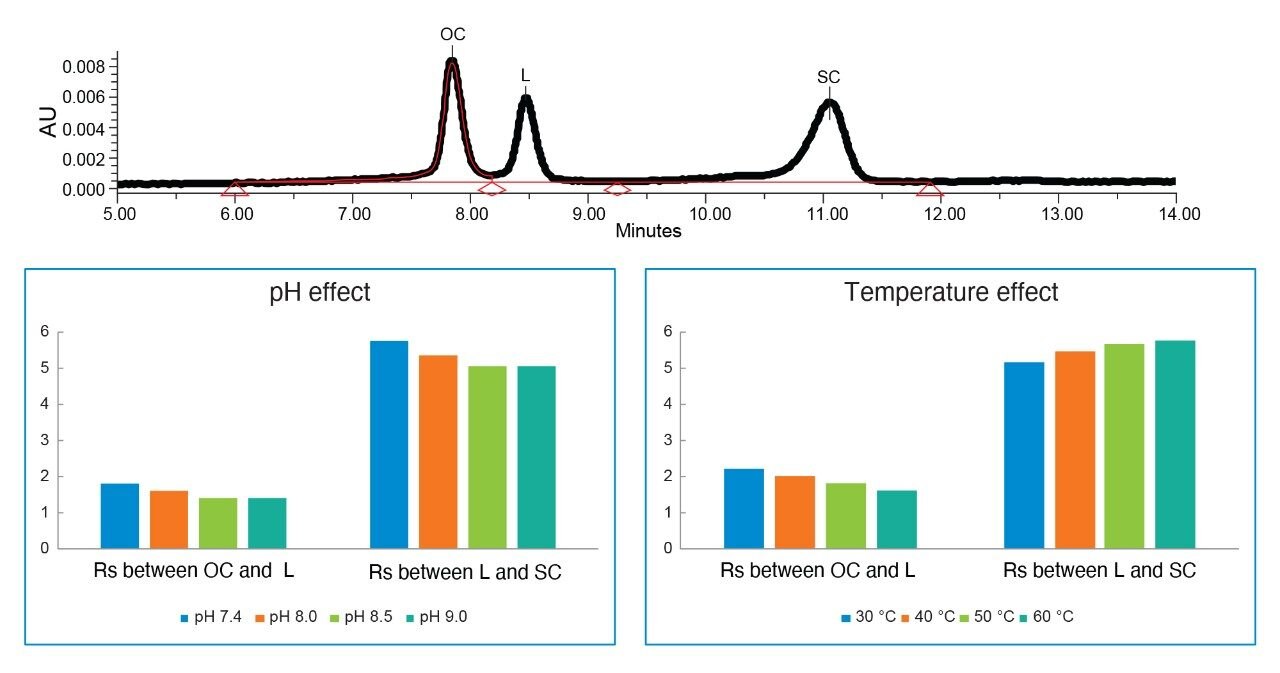
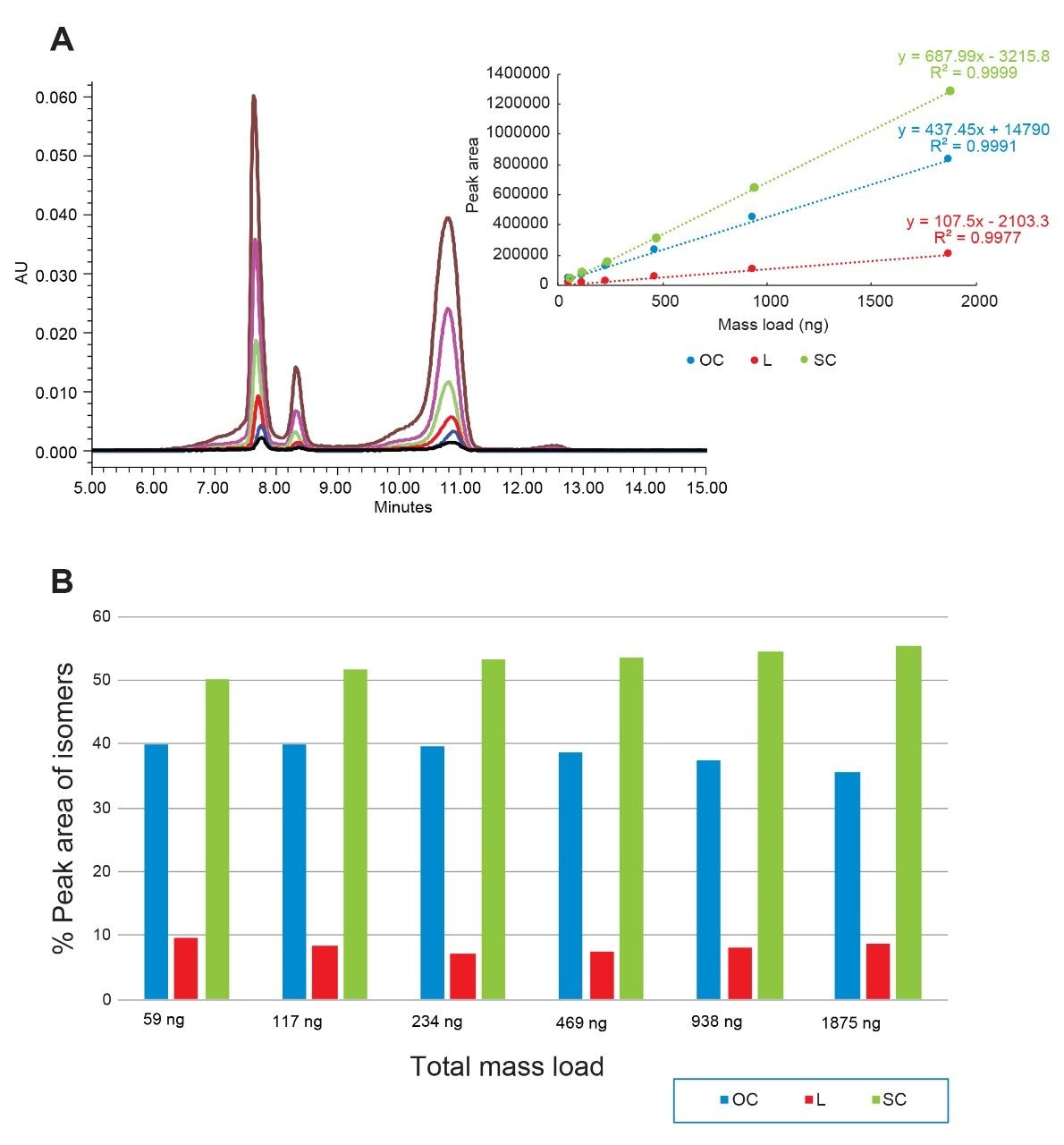
Plasmids are small circular double-stranded DNAs and have been increasingly used in the manufacturing of gene therapies and vaccines in recent years. The production of plasmid DNA mostly results in the supercoiled form, which is the desired conformation due to its excellent stability and favorable antigenicity. However, other forms of plasmid, such as open circular and linear forms, are usually present due to conformational changes that occur during bioprocessing. The open circular form results when the plasmid is nicked on one strand, while the linear form is produced when the plasmid is nicked on both strands at or near the same site. These isoforms, along with others, are considered plasmid impurities, because they may affect the efficacy or the safety of the products.1,2 For this reason, FDA guidelines recommend that the purity of the supercoiled form of the plasmid be greater than 80%.2,3 To ensure high purity and quality, the industry expectation for the supercoiled plasmid purity level is usually greater than 90%.2,3 Consequently, reliable measurement on the relative abundance of plasmid isoforms becomes critical to ensure batch process, formulation, and storage consistency.
For decades, the use of agarose gel electrophoresis has been considered as routine analytical method for separating plasmid isoforms.4,5 However, this technique has drawbacks such as difficulty in quantification, low throughput, and manual-handling steps. Capillary electrophoresis (CE) has also been used for plasmid isoform analysis,4 but it struggles to analyze samples that are in the complex matrix such as cell lysate. Anion-exchange chromatography (AEX) separates analytes based on their differences in the amount and localization of negative surface charges. It is well accepted that AEX is a robust, reproducible, easily automated analytical technique, requiring small amounts of samples. Since plasmid DNAs are negatively charged due to the phosphate groups on the backbone, AEX is a natural choice for plasmid-related separation and analysis.4,5
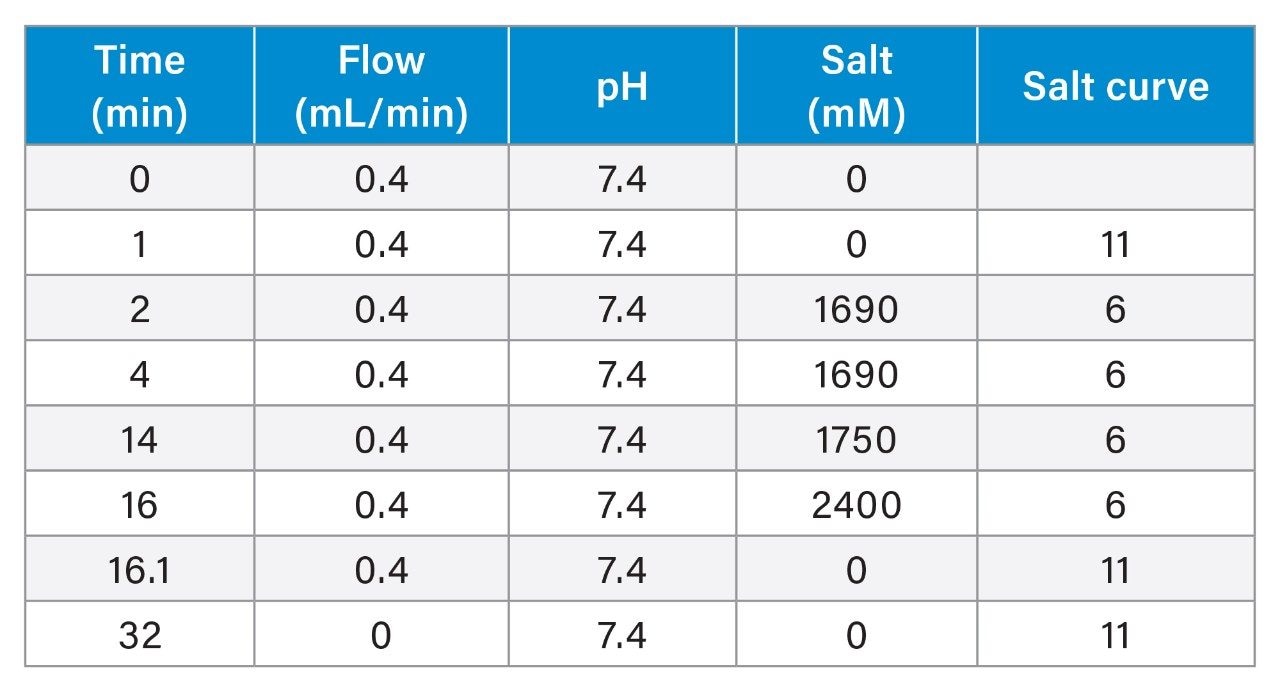
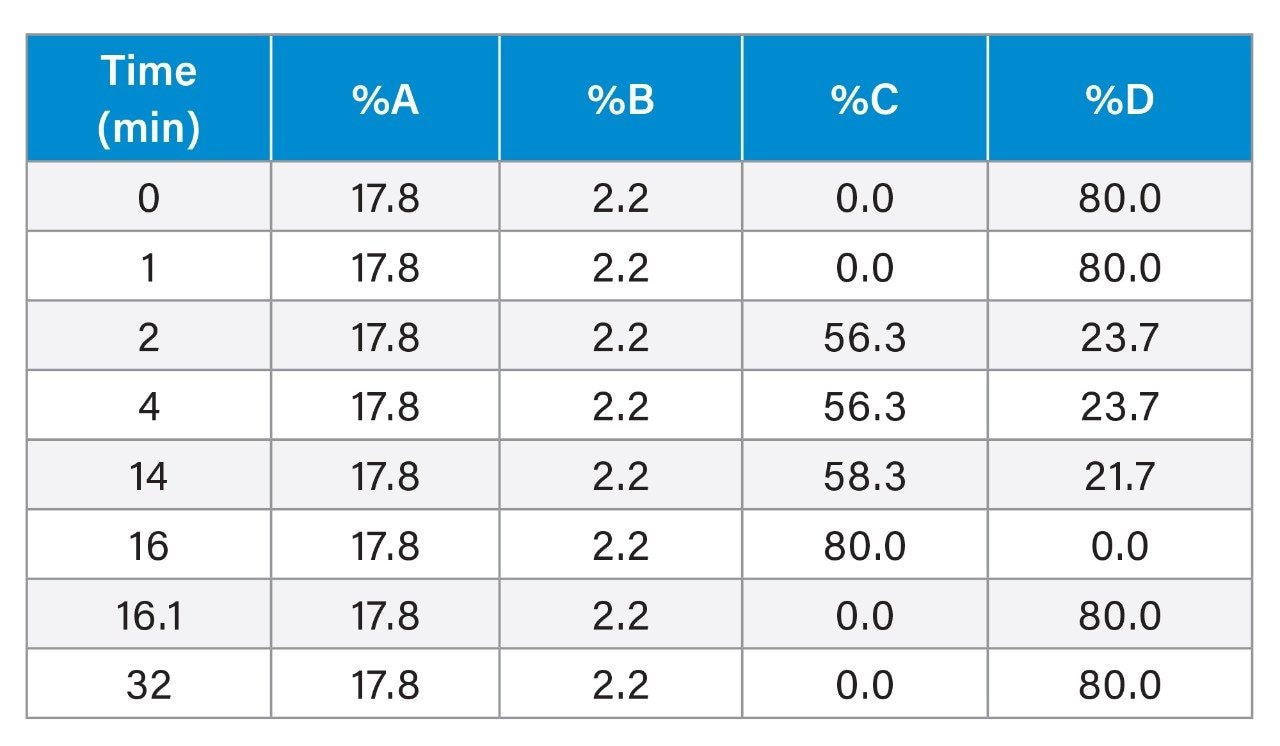
In this application note, we show that various levels of the three major isoforms of the plasmid DNA, namely, supercoiled, open circular, and linear form, can be separated and quantified on a Waters Protein-Pak Hi Res Q Column fitted to an ACQUITY UPLC H-Class Bio System.